cyklus glukóza-alanin, nebo Cahill cyklu, navrhl poprvé do Mallette, Exton a Park, a Felig et al. mezi lety 1969 a 1970, se skládá z řady kroků, jimiž extrahepatálních tkáních, např. v kosterním svalu, export pyruvát a amino skupiny jako alanin jater, a přijímat glukózu z jater prostřednictvím krevního řečiště.
hlavní kroky cyklu glukóza-alanin jsou shrnuty níže.
- když se v extrahepatálních tkáních používají aminokyseliny pro energii, pyruvát, odvozený od glykolýzy, se používá jako akceptor aminoskupiny, tvořící alanin, neesenciální aminokyselinu.
- alanin difunduje do krevního řečiště a dosáhne jater.
- v játrech se aminoskupina alaninu přenáší na α-ketoglutarát za vzniku pyruvátu a glutamátu.
- aminoskupina glutamátu většinou vstupuje do močovinového cyklu a částečně působí jako donor dusíku v mnoha biosyntetických drahách.
pyruvát vstupuje do glukoneogeneze a používá se k syntéze glukózy. - nově vytvořená glukóza difunduje do krevního řečiště a dostává se do periferních tkání, kde se v důsledku glykolýzy převede na pyruvát, který může přijímat aminoskupiny z volných aminokyselin, čímž uzavírá cyklus.
proto cyklus glukóza-alanin poskytuje vazbu mezi metabolismem sacharidů a aminokyselin, jak je schematicky popsáno níže.
Glukosa → Pyruvát → Alanin → Pyruvát → Glukóza

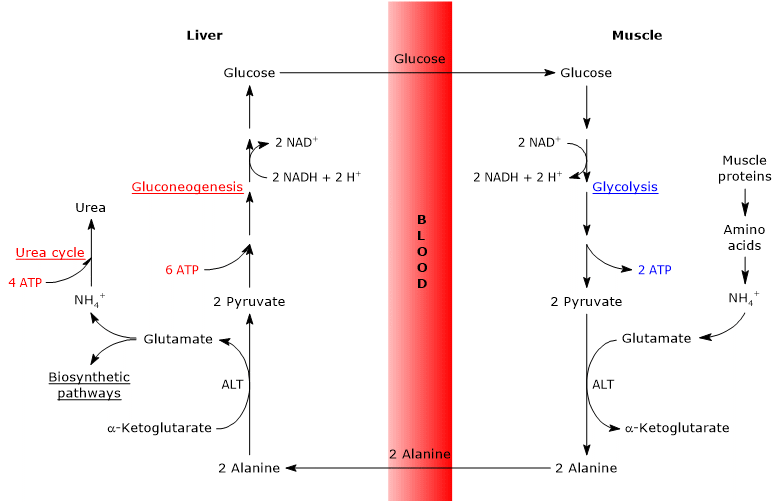
cyklus glukóza-alanin se vyskytuje nejen mezi kosterního svalstva, první tkáně, ve které bylo zjištěno, a jater, ale zahrnuje jiné buňky a extrahepatálních tkání včetně buněk imunitního systému, jako jsou lymfatické orgány.
OBSAH
- kroky cyklus glukóza-alanin
- Cahill cyklu v kosterním svalu
- Cahill cyklus v játrech
- Transamináz
- Funkce cyklus glukóza-alanin
- náklady na Energie glukóza-alaninový cyklus
- Podobnosti a rozdíly mezi Cahill cyklus a Coriho cyklu
kroky cyklus glukóza-alanin
analýza kroků cyklus glukóza-alanin je vyroben s ohledem na cyklus mezi kosterní sval a játra.
jak intracelulární, tak extracelulární proteiny jsou kontinuálně hydrolyzovány na aminokyseliny, které tvoří, a resyntetizovány, a rychlost, při které se tyto procesy vyskytují, je přesně vyvážená, čímž se zabrání ztrátě hmotnosti bez tuku.
avšak za katabolických podmínek, jako je intenzivní a prodloužené cvičení nebo půst, rychlost rozpadu svalových bílkovin překračuje syntézu. To vede k uvolnění aminokyselin, z nichž některé se používají pro energii a jiné pro glukoneogenezi. A oxidaci uhlíkatých koster aminokyselin, zejména aminokyselin s rozvětveným řetězcem nebo BCAA (leucin, isoleucin a valin), může být významným zdrojem energie pro svaly. Například po asi 90 minutách namáhavého cvičení poskytuje oxidace aminokyselin ve svalech 10-15% energie potřebné pro kontrakci.
využití uhlíkatých koster aminokyselin pro energetické zahrnuje odstranění aminoskupiny, a pak vylučování aminodusíku v netoxické formě.
odstranění α-aminoskupiny dochází prostřednictvím transaminace, které lze shrnout takto:
α-Keto kyseliny + Aminokyseliny ⇄ Nové aminokyseliny + Nové α-keto kyseliny
Takové reakce, katalyzované enzymy, tzv. aminotransferázy nebo transamináz (ES 2.6.1) jsou volně reverzibilní.
aminokyselin s Rozvětveným řetězcem, například přenos aminoskupiny na α-ketoglutarát nebo 2-oxoglutaric kyseliny, tvoří glutamát a α-keto kyseliny odvozené od původní aminokyselina, v reakci katalyzované rozvětveným řetězcem aminotransferázy nebo BCAT (ES 2.6 .1.42).
Cahill cyklu v kosterním svalu
V kosterním svalu, nově vytvořené glutamátu může reagovat s amoniakem za vzniku glutaminu, pro mnoho tkání a orgánů, jako je mozek, hlavní vozidla pro meziorgánových propojení metabolických transport dusíku. Reakce je katalyzována cytosolickým enzymem glutamin syntetázou (EC 6.3.1.2) a spotřebovává ATP.
Glutamát + NH4+ + ATP → Glutamin + ADP + Pi
V tomto případě, glutamát opustí Cahill cyklu.
Alternativně, a v kontrastu k tomu, co se děje ve většině ostatních tkání, nově vytvořené glutamátu může převést aminokyselin skupiny pyruvátu, odvozený od glykolýza, tvoří alanin a α-ketoglutarát. Tato transaminace je katalyzována alaninaminotransferázou nebo ALT (EC 2.6.1.2), což je enzym nacházející se ve většině živočišných a rostlinných tkání.
Glutamát + Pyruvát ⇄ Alanin + α-Ketoglutarát
alanin vyrábí a které pocházejí přímo z odbourávání bílkovin a svalové bílkoviny jsou bohaté na alanin, může opustit buňku a být nesen krevním řečištěm do jater; v tomto způsob, amino skupina dosáhne jater. A rychlost přenosu alaninu vytvořeného transaminací pyruvátu do oběhu je úměrná produkci intracelulárního pyruvátu.
Poznámka: alanin a glutamin jsou hlavními zdroji dusíku a uhlíku v metabolismu aminokyselin mezi organismy.
Cahill cyklus v játrech
Poté, co v játrech, jaterní alanin aminotransferázy katalyzuje transaminace, ve které alanin, hlavní gluconeogenic aminokyselin, působí jako amino skupiny dárce a α-ketoglutarát jako α-keto kyseliny akceptor. Produkty reakce jsou pyruvát, tj. uhlíková kostra alaninu a glutamát.
Alanin + α-Ketoglutarát ⇄ Glutamát + Pyruvát
Glutamát, v reakci katalyzované glutamát dehydrogenázy (ES 1.4.1.2), enzym přítomný v mitochondriální matrici, tvoří amonný iont, který vstupuje do močovinového cyklu, a α-ketoglutarát, který může vstoupit do Krebsova cyklu. Tato reakce je anaplerotická reakce, která spojuje metabolismus aminokyselin s krebsovým cyklem.
Glutamát + H2O + NAD+ ⇄ α-Ketoglutarát + NH4+ + NADH + H+
Nicméně, glutamát může také reagovat s oxalacetátu na formě aspartátu a α-ketoglutarát, v reakci katalyzované aspartátaminotransferázy (EC 2.6.1.1). Aspartát se podílí na tvorbě močoviny, stejně jako na syntéze purinů a pyrimidinů.
Glutamát + Oxalacetát ⇄ Aspartát + α-Ketoglutarát
Také poté, co pyruvát vznikne mohou mít různé metabolické osudy: to může být oxidovány za produkce ATP, a pak odejít cyklus glukóza-alanin, nebo zadejte glukoneogeneze cestu, a tak pokračovat v cyklu.
produkovaná glukóza se uvolňuje z jater do krevního řečiště a dodává se do různých tkání, které ji vyžadují, jako kosterní sval, ve kterém se používá pro syntézu pyruvátu. Nově vytvořený pyruvát může zase reagovat s glutamátem, čímž uzavírá cyklus.
Transaminázy
Jak již bylo zmíněno, odstranění aminoskupiny z aminokyselin dochází prostřednictvím transaminace (viz výše pro obecné reakce). Tyto reakce jsou katalyzovány enzymy nazývanými aminotransferázy nebo transaminázy.
jsou cytosolové enzymy, přítomné ve všech buňkách a obzvláště hojně se vyskytuje v játrech, ledvinách, střeva a svalů; vyžadují pyridoxalfosfát nebo PLP, aktivní forma vitamínu B6, nebo pyridoxin, jako koenzym, který je pevně vázán na aktivní místo.
V transaminační reakcí amino skupiny volných aminokyselin, s výjimkou threonin a lysin, je použito na malý počet α-keto kyseliny, především pyruvát, oxalacetát a α-ketoglutarát.
buňky obsahují různé typy aminotransferáz: mnohé jsou specifické pro α-ketoglutarát jako akceptor kyseliny α-keto, ale liší se specificitou pro aminokyselinu, ze které jsou pojmenovány. Příklady jsou výše zmíněné alaninaminotransferázy, také volal alanin transaminázy a glutamové pyrohroznové transferázy nebo GPT, a aspartátaminotransferázy nebo AST, také volal glutamát-oxaloctová transamináza nebo.
je třeba zdůraznit, že při těchto reakcích nedochází k čisté deaminaci, ke ztrátě aminoskupin, protože akceptor α-keto kyseliny je aminován a aminokyselina deaminována.
funkce glukózo-alaninového cyklu
tento cyklus má různé funkce.
- transportuje dusík v netoxické formě z periferních tkání do jater.
- transportuje pyruvát, glukoneogenní substrát, do jater.
- odstraňuje pyruvát z periferních tkání. To vede k vyšší produkci ATP z glukózy v těchto tkáních. Ve skutečnosti může NADH produkovaný během glykolýzy vstoupit do mitochondrií a být oxidován oxidační fosforylací.
- umožňuje udržovat relativně vysokou koncentraci alaninu v hepatocytech, dostatečnou k inhibici degradace bílkovin.
- může hrát roli v obraně hostitele proti infekčním chorobám.
konečně je důležité zdůraznit, že v cyklu glukóza-alanin neexistuje žádná čistá syntéza glukózy.
energetické náklady cyklu glukóza-alanin
stejně jako cyklus Cori má také cyklus glukóza-alanin náklady na energii rovnající se 3-5 ATP.
část cyklu, která probíhá v periferních tkáních zahrnuje výrobu 5-7 ATP na jednu molekulu glukózy:
- 2 ATP je produkován glykolýza;
- 3-5 ATP odvodit z NADH/FADH2 (viz níže).
Místo toho, v játrech, v játrech a v cyklu močoviny náklady 10 ATP:
- 6 ATP jsou spotřebované v průběhu glukoneogeneze na jednu molekulu glukózy syntetizovány;
- 4 ATP jsou spotřebované v cyklu močoviny v molekule močoviny syntetizován.
cyklus glukózy a alaninu, stejně jako cyklus Cori, přesouvá část metabolické zátěže z extrahepatálních tkání do jater. Nicméně, náklady na energie hradí jater je odůvodněno výhody, že cyklus přináší na celé tělo, jako to umožňuje, zejména podmínky, efektivní rozklad bílkovin v extrahepatálních tkáních (zejména v kosterních svalech), což umožňuje získat gluconeogenic substráty, stejně jako použití aminokyselin pro energetické v extrahepatálních tkáních.
podobnosti a rozdíly mezi cyklem Cahill a cyklem Cori
existují některé analogie mezi oběma cykly, které jsou uvedeny níže.
- Cahill cyklu částečně překrývá Cori cyklu, kdy pyruvát je přeměněn na glukózu a monosacharid je transportován do extrahepatálních tkání, ve kterém je převeden opět na pyruvát přes glycolytic pathway.
- vstup do dráhy glukoneogeneze je pro oba cykly podobný: alanin i laktát se převádějí na pyruvát.
- Jako Coriho cyklus, cyklus glukóza-alanin se vyskytuje mezi různými typy buněk, na rozdíl od metabolických drah, jako jsou glykolýza, Krebsův cyklus, nebo glukoneogeneze, které se vyskytují v rámci jednotlivých buněk

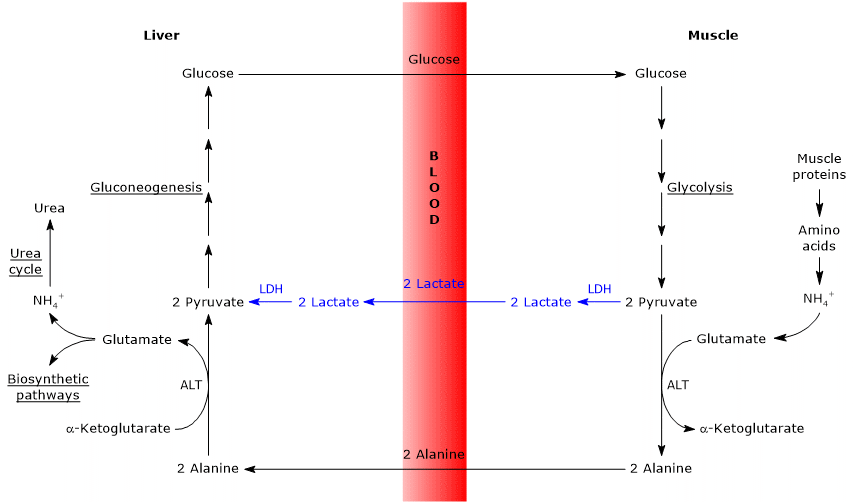
Níže některé rozdíly mezi dvěma cykly.
- hlavní rozdíl se týká tří uhlíkových meziproduktů, které se z periferních tkání dostávají do jater: laktát v cyklu Cori a alanin v cyklu glukóza-alanin.
- další rozdíl se týká osudu NADH produkovaného glykolýzou v periferních tkáních.
V Coriho cyklu, koenzym působí jako redukční činidlo pro snížení pyruvát na laktát, v reakci katalyzované laktát dehydrogenáza (EC 1.1.1.27).
cyklus glukóza-alanin, tato redukce se nevyskytuje, a elektrony z NADH + h + může být transportován do mitochondrie přes malát-aspartát a glycerol-3-fosfát raketoplány, vytváří NADH, první raketoplán, a FADH2, druhý raketoplán. A výtěžek ATP z NADH a FADH2 je 2,5 a 1,5. - konečně z předchozího bodu je zřejmé, že na rozdíl od cyklu Cori vyžaduje cahillův cyklus přítomnost kyslíku a mitochondrií v periferních tkáních.
Berg J. M., Tymoczko jl, Stryer L. Biochemie. 5. vydání. W. h. Freeman a společnost, 2002
Felig P., Pozefsk T., Marlis e., Cahill G. F. Alanine: klíčová role v glukoneogenezi. Věda 1970; 167 (3920): 1003-4. doi: 10.1126 / věda.167.3920.1003
Gropper. S. S., Smith J. L., Groff J. L. Pokročilá výživa a metabolismus člověka. Cengage Learning, 2009
Lecker. S. H., Goldberg A. L. a Mitch W. E. degradace Proteinů pomocí ubiquitin–proteazomové dráhy v normální a chorobné stavy. J Am Soc Nephrol 2006; 17 (7): 1807-19. doi: 10.1681 / ASN.2006010083
Mallette L. E., Exton J. H., a Park C. R. Ovládání glukoneogeneze z aminokyselin v perfundovaných játrech potkanů. J Biol Chem 1969; 244 (20): 5713-23.
Nelson D. L., Cox M. M. Lehninger. Principy biochemie. 6. vydání. W. h. Freeman and Company, 2012
Raju S. M., Madala B. Ilustrovaná lékařská biochemie. Jaypee Brothers Publishers, 2005
Wu G. aminokyseliny: biochemie a výživa. CRC Press, 2013