glukoosi-alaniinisykli eli Cahill-sykli, jota Mallette, Exton ja Park sekä Felig et al ehdottivat ensimmäistä kertaa. vuosina 1969-1970 se koostuu vaiheista, joiden kautta ekstrahepaattiset kudokset, esimerkiksi luurankolihas, vievät pyruvaatti-ja aminoryhmiä alaniinina maksaan ja saavat glukoosia maksasta verenkierron kautta.
alla on yhteenveto glukoosi-alaniinisyklin tärkeimmistä vaiheista.
- kun ekstrahepaattisissa kudoksissa käytetään aminohappoja energiaksi, glykolyysistä peräisin olevaa pyruvaattia käytetään aminoryhmän hyväksyjänä muodostaen alaniinia, joka on epäolennainen aminohappo.
- alaniini diffundoituu verenkiertoon ja päätyy maksaan.
- maksassa alaniinin aminoryhmä siirtyy α-ketoglutaraatiksi muodostaen pyruvaattia ja glutamaattia.
- glutamaatin aminoryhmä siirtyy pääosin ureakiertoon ja toimii osittain typen luovuttajana monissa biosynteettisissä reiteissä.
pyruvaatti pääsee glukoneogeneesiin ja sitä käytetään glukoosisynteesissä. - vastamuodostunut glukoosi diffundoituu verenkiertoon ja saavuttaa perifeeriset kudokset, joissa glykolyysin seurauksena muuttuu pyruvaatiksi, joka voi hyväksyä vapaiden aminohappojen aminoryhmiä, jolloin kierto sulkeutuu.
glukoosi-alaniinisykli muodostaa näin ollen hiilihydraatti-ja aminohappoaineenvaihdunnan välisen yhteyden, kuten alla kuvataan.
glukoosi → pyruvaatti → alaniini → pyruvaatti → glukoosi

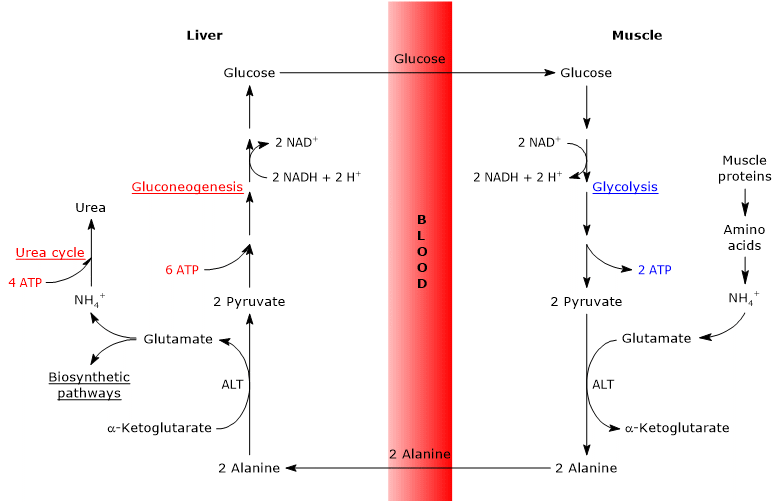
glukoosi-alaniinisykli ei tapahdu ainoastaan luustolihaksen, ensimmäisen kudoksen, jossa se havaittiin, ja maksan välillä, vaan siihen liittyy muita soluja ja ekstrahepaattisia kudoksia, mukaan lukien immuunijärjestelmän soluja, kuten imukudoselimiä.
sisältö
- glukoosi-alaniinisyklin vaiheet
- Cahill-sykli luurankolihaksissa
- Cahill-sykli maksassa
- transaminaasit
- glukoosi-alaniinisyklin toiminnot
- glukoosi-alaniinisyklin energiakustannukset
- Cahill-ja Cori-syklien yhtäläisyydet ja erot
glukoosi-alaniinisyklin vaiheet
glukoosi-alaniinisyklin vaiheet analysoidaan ottaen huomioon luustolihasten ja maksan välinen sykli.
sekä solunsisäiset että solunulkoiset proteiinit hydrolysoituvat jatkuvasti aminohapoiksi ja resyntetisoituvat, ja näiden prosessien nopeus on tasapainossa, mikä estää rasvattoman massan häviämisen.
kuitenkin katabolisissa olosuhteissa, kuten intensiivisessä ja pitkäkestoisessa liikunnassa tai paastossa, lihasproteiinin hajoamisnopeus ylittää synteesin. Tämä johtaa aminohappojen vapautumiseen, joista osaa käytetään energiaan ja toisia glukoneogeneesiin. Ja aminohappojen, erityisesti haaraketjuisten aminohappojen tai BCAA: n (leusiini, isoleusiini ja valiini) hiilirungon hapettuminen voi olla merkittävä energianlähde lihakselle. Esimerkiksi noin 90 minuutin rasittavan liikunnan jälkeen aminohapon hapettuminen lihaksessa tuottaa 10-15% supistumiseen tarvittavasta energiasta.
aminohappojen hiilirungon hyödyntäminen energiaksi edellyttää aminoryhmän poistamista ja sen jälkeen aminotypen erittymistä myrkyttömässä muodossa.
α-aminoryhmä poistuu transaminaatiolla, mikä voidaan tiivistää seuraavasti:
α-ketohappo + aminohappo ⇄ Uusi aminohappo + Uusi α-ketohappo
tällaiset aminotransferaasi-tai transaminaasientsyymien (EC 2.6.1) katalysoimat reaktiot ovat vapaasti palautuvia.
haaraketjuiset aminohapot esimerkiksi siirtävät aminoryhmän α-ketoglutaraatiksi tai 2-oksoglutaarihapoksi, jolloin muodostuu glutamaattia ja alkuperäisestä aminohaposta johdettua α-ketohappoa haaraketjuaminotransferaasin tai BCAT: n katalysoimassa reaktiossa (EC 2.6 .1.42).
Cahill-sykli luurankolihaksissa
luurankolihaksissa vastamuodostunut glutamaatti voi reagoida ammoniakin kanssa muodostaen glutamiinia monille kudoksille ja elimille, kuten aivoille, jotka ovat tärkein typen kuljetusväline interorgaanisessa kuljetuksessa. Reaktiota katalysoi sytosolikoentsyymi glutamiinisyntetaasi (EC 6.3.1.2) ja kuluttaa ATP: tä.
glutamaatti + NH4+ + ATP → glutamiini + ADP + Pi
tällöin glutamaatti poistuu Cahill-syklistä.
Vaihtoehtoisesti ja toisin kuin useimmissa muissa kudoksissa, vasta muodostunut glutamaatti voi siirtää aminoryhmän glykolyysissä johdetulle pyruvaatille muodostaen alaniinia ja α-ketoglutaraattia. Tätä transaminaatiota katalysoi alaniiniaminotransferaasi tai ALAT (EC 2.6.1.2), jota esiintyy useimmissa eläin-ja kasvikudoksissa.
glutamaatti + pyruvaatti ⇄ alaniini + α-ketoglutaraatti
tuotettu ja suoraan proteiinin hajoamisesta saatu alaniini ja lihasproteiinit, joissa on runsaasti alaniinia, voivat poistua solusta ja kulkeutua verenkiertoon maksaan; näin aminoryhmä pääsee maksaan. Ja nopeus, jolla pyruvaatin transaminaatiossa muodostuva alaniini siirtyy kiertoon, on verrannollinen pyruvaatin solunsisäiseen tuotantoon.
Huom.alaniini ja glutamiini ovat tärkeimmät typen ja hiilen lähteet interorgaaniaminohapon aineenvaihdunnassa.
Cahill-sykli maksassa
kerran maksassa maksan alaniiniaminotransferaasi katalysoi transaminaatiota, jossa pääasiallinen glukoneogeeninen aminohappo alaniini toimii aminoryhmän luovuttajana ja α-ketoglutaraatti α-ketohapon hyväksyjänä. Reaktion tuotteita ovat pyruvaatti eli alaniinin hiilirunko ja glutamaatti.
alaniini + α-ketoglutaraatti ⇄ glutamaatti + pyruvaatti
glutamaatti glutamaattidehydrogenaasin katalysoimassa reaktiossa (EC 1.4.1.2), mitokondrio-matriisissa esiintyvä entsyymi, muodostaa ammoniumionin, joka menee ureakiertoon, ja α-ketoglutaraatin, joka voi päästä Krebs-kiertoon. Tämä reaktio on anapleroottinen reaktio, joka yhdistää aminohappoaineenvaihdunnan Krebs-sykliin.
glutamaatti + H2O + nad+ ⇄ α-ketoglutaraatti + NH4+ + NADH + H+
glutamaatti voi kuitenkin reagoida myös oksaloasetaatin kanssa muodostaen aspartaattia ja α-ketoglutaraattia aspartaattiaminotransferaasin (EC 2.6.1.1) katalysoimassa reaktiossa. Aspartaatti osallistuu urean muodostumiseen sekä puriinien ja pyrimidiinien synteesiin.
glutamaatti + Oksaloasetaatti ⇄ aspartaatti + α-ketoglutaraatti
myös tuotetulla pyruvaatilla voi olla erilaisia metabolisia kohtaloita: se voi hapettua ATP: n tuotantoa varten, minkä jälkeen se poistuu glukoosi-alaniinisyklistä tai siirtyy glukoneogeneesireittiin ja siten jatkaa syklissä.
tuotettu glukoosi vapautuu maksasta verenkiertoon ja kulkeutuu sitä tarvitseviin eri kudoksiin, kuten luurankolihakseen, jossa sitä käytetään pyruvaattisynteesissä. Vastamuodostunut pyruvaatti voi puolestaan reagoida glutamaatin kanssa, jolloin sykli sulkeutuu.
transaminaasit
kuten aiemmin mainittiin, aminoryhmä poistuu aminohapoista transaminaation kautta (katso yllä oleva yleisreaktio). Näitä reaktioita katalysoivat aminotransferaasi-tai transaminaasientsyymit.
ne ovat sytosolientsyymejä, joita on kaikissa soluissa ja erityisen runsaasti maksassa, munuaisissa, suolistossa ja lihaksissa; ne tarvitsevat koentsyymikseen pyridoksaalifosfaattia tai PLP: tä, B6-vitamiinin tai pyridoksiinin aktiivista muotoa, joka on tiukasti sitoutuneena aktiiviseen kohtaan.
transaminaatioreaktioissa vapaiden aminohappojen aminoryhmä treoniinia ja lysiiniä lukuun ottamatta kanavoituu muutamille α-ketohapoille, erityisesti pyruvaatille, oksaloasetaatille ja α-ketoglutaraatille.
solut sisältävät erityyppisiä aminotransferaaseja: monet ovat spesifisiä α-ketoglutaraatille α-ketohapon hyväksyjänä, mutta eroavat spesifisyydeltään aminohapolle, josta ne on nimetty. Esimerkkejä ovat edellä mainittu alaniiniaminotransferaasi, jota kutsutaan myös alaniinitransaminaasiksi ja glutamiinipyruviinitransferaasiksi tai GPT: ksi, ja aspartaattiaminotransferaasi tai ASAT, jota kutsutaan myös glutamiinioksaloetikkatransaminaasiksi tai GOT: ksi.
on korostettava, että näissä reaktioissa ei tapahdu nettodesaminaatiota eikä aminoryhmien häviämistä, koska α-ketohapon hyväksyjä aminoituu ja aminohappo deaminoituu.
glukoosi-alaniinisyklin tehtävät
tällä syklillä on useita tehtäviä.
- se kuljettaa typpeä myrkyttömässä muodossa perifeerisistä kudoksista maksaan.
- se kuljettaa pyruvaattia, glukoneogeenista substraattia, maksaan.
- se poistaa pyruvaatin perifeerisistä kudoksista. Tämä johtaa ATP: n suurempaan tuotantoon glukoosista näissä kudoksissa. Glykolyysin aikana syntyvä NADH voi nimittäin päästä mitokondrioihin ja hapettua oksidatiivisen fosforylaation kautta.
- se mahdollistaa suhteellisen korkean alaniinipitoisuuden ylläpitämisen maksasoluissa, mikä riittää estämään proteiinin hajoamista.
- sillä voi olla merkitystä isäntien puolustautumisessa tartuntatauteja vastaan.
lopuksi on tärkeää korostaa, ettei glukoosin nettosynteesiä tapahdu glukoosi-alaniinisyklissä.
glukoosi-alaniinisyklin energiakustannukset
Cori-syklin tapaan myös glukoosi-alaniinisyklin energiakustannukset ovat 3-5 ATP.
se syklin osa, joka tapahtuu perifeerisissä kudoksissa, sisältää 5-7 ATP: tä glukoosimolekyyliä kohti.:
- 2 ATP: tä tuotetaan glykolyysillä;
- 3-5 ATP: tä saadaan NADH: sta/FADH2: sta (KS.alla).
sen sijaan maksassa glukoneogeneesi ja ureakierto maksavat 10 ATP: tä:
- 6 ATP: tä kulutetaan glukoneogeneesin aikana syntetisoitua glukoosimolekyyliä kohti;
- 4 ATP: tä kulutetaan ureakierrossa syntetisoitua ureamolekyyliä kohti.
glukoosi-alaniinisykli, kuten Cori-syklikin, siirtää osan metabolisesta rasituksesta ekstrahepaattisista kudoksista maksaan. Maksan maksamaa energiakustannusta voidaan kuitenkin perustella syklin koko keholle tuomilla eduilla, sillä se mahdollistaa tietyissä olosuhteissa proteiinien tehokkaan jakautumisen ekstrahepaattisissa kudoksissa (erityisesti luurankolihaksissa), mikä puolestaan mahdollistaa glukoneogeenisten substraattien saamisen sekä aminohappojen käytön energiaksi ekstrahepaattisissa kudoksissa.
yhtäläisyyksiä ja eroja Cahillin ja Corin syklin välillä
näiden kahden syklin välillä on joitakin analogioita, jotka on lueteltu alla.
- Cahill-sykli menee osittain päällekkäin Cori-syklin kanssa, kun pyruvaatti muuttuu glukoosiksi ja monosakkaridi kulkeutuu ekstrahepaattisiin kudoksiin, joissa se muuntuu glykolyyttistä reittiä uudelleen pyruvaatiksi.
- glukoneogeneesin reitti on samanlainen kahden syklin ajan: sekä alaniini että laktaatti muuttuvat pyruvaatiksi.
- Cori-syklin tavoin glukoosi-alaniinisykli tapahtuu eri solutyyppien välillä, toisin kuin yksittäisissä soluissa tapahtuvat aineenvaihduntareitit, kuten glykolyysi, Krebs-sykli tai glukoneogeneesi

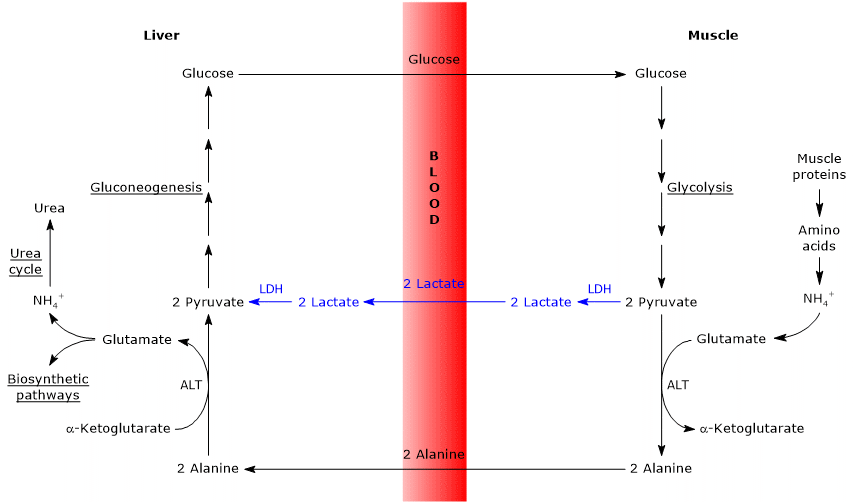
alla joitakin eroja näiden kahden syklin välillä.
- suurin ero koskee kolmen hiilen välituotetta, joka perifeerisistä kudoksista päätyy maksaan: laktaattia Cori-syklissä ja alaniinia glukoosi-alaniinisyklissä.
- toinen ero koskee glykolyysissä syntyvän NADH: n kohtaloa perifeerisissä kudoksissa.
Cori-syklissä koentsyymi toimii pelkistimenä pelkistäen pyruvaatin laktaatiksi laktaattidehydrogenaasin katalysoimassa reaktiossa (EC 1.1.1.27).
glukoosi-alaniinisyklissä tätä pelkistymistä ei tapahdu ja NADH: n elektronit voivat kulkeutua mitokondrioihin malaatti-aspartaatti-ja glyseroli-3-fosfaattisukkuloiden kautta, jolloin syntyy NADH, ensimmäinen sukkula, ja fadh2, toinen sukkula. ATP: n tuotto NADH: lta ja FADH2: lta on vastaavasti 2,5 ja 1,5. - lopulta edellisestä pisteestä on selvää, että toisin kuin Cori-sykli, Cahill-sykli vaatii hapen ja mitokondrioiden läsnäoloa perifeerisissä kudoksissa.
Berg J. M., Tymoczko J. L., and Stryer L. Biochemistry. 5. Painos. W. H. Freeman and Company, 2002
Felig P., Pozefsk T., Marlis E., Cahill G. F. Alanine: key role in gluconeogenesis. Science 1970;167(3920): 1003-4. doi: 10.1126 / tiede.167. 3920. 1003
Gropper S. S., Smith J. L., Groff J. L. Kehittynyt ravitsemus ja ihmisen aineenvaihdunta. Cengage Learning, 2009
Lecker S. H., Goldberg A. L. and Mitch W. E. Protein degradation by the ubikitin–proteasome pathage in normal and disease states. J Am Soc Nefrol 2006; 17(7): 1807-19. doi: 10.1681 / ASN.2006010083
Mallette L. E., Exton J. H. ja Park C. R. Control of glukoneogenesis from aminohapot in the perfused rotan maksa. J Biol Chem 1969;244 (20):5713-23.
Nelson D. L., Cox M. M. Lehninger. Biokemian periaatteet. 6. Painos. W. H. Freeman and Company, 2012
Raju S. M., Madala B. Kuvitettu lääketieteellinen biokemia. Jaypee Brothers Publishers, 2005
Wu G. Amino acids: biochemistry and nutrition. CRC Press, 2013