広告:
この記事では、ユーグレナViridisについて説明します:-1。 ユーグレナ・ヴィリディスの生息環境と生息地2. ユーグレナ・ヴィリディスの培養3. 構造4. 第5話では、”機動戦士ガンダム 栄養6. 7. 排泄8. 行動9. 再生産10. ポジションは11。 他のいくつかのユーグレノイド鞭毛虫。
:
- ミドリイシの習慣と生息地
- ミドリイシの文化
- ミドリイシの構造
- ミドリイシの移動
- ミドリイシの栄養
- ミドリイシの呼吸
- ミドリムシの排泄
- ミドリムシの行動
- ミドリムシの繁殖
- ミドリムシの位置
- その他のミドリムシ鞭毛虫
1. ユーグレナグリーンの習慣と生息地:
ユーグレナグリーン(Gr. eu=真; glene=目球または目瞳孔;L.、viridis=緑)は、一般的で孤独で自由な生きている淡水鞭毛虫です。 それは淡水のプール、池、溝およびゆっくり動く流れにあります。 それは植生のかなりの量がある豊富に発見されます。
:
小枝、葉、動物の顔など、腐敗する窒素性有機物を含むよく維持された庭園の池。、この生物の良い源です。 それは一般的に属の他の種と一緒に住んでいます。 それらは時々水に明瞭な緑がかった色を与えるか、または池水の表面の浮きかすの緑のフィルムを形作ることほど多数である。
2. ユーグレナビリディスの培養:
ユーグレナビリディスの培養は、以下の方法により実験室で容易に調製することができる。 牛や馬の糞を蒸留水で半開きにして2日間冷やします。 その後、ミドリムシを含む池からの雑草を瓶に入れ、明るい窓の近くに瓶を置きます。 数日後に、この窒素注入にユーグレナが現れるでしょう。
3. ミドリムシの構造:
形状:
広告:
Euglena viridisは細長く、外観は紡錘形である。 前端は鈍く、中央部はより広く、後端は尖っている。
サイズ:
ユーグレナビリディスは、体の最も厚い部分で長さが約40-60ミクロン、幅が14-20ミクロンです。
ペリクル:
体は薄く、柔軟で、丈夫で強いクチクラペリプラストまたはペリクルで覆われており、原形質膜の下にあります。 それはすべてのラウンドmyonemesと呼ばれる斜めが平行な線条を持っています。 しかし、Chadefaud(1937)によると、ペリクルは外側の薄い層のエピキュティクルと内側の厚い層のキューティクルでできています。
ペリクルの両方の層は体のいたるところに存在しますが、epicuticleのみが前方に配置された細胞咽頭と貯水池に終わります。
ペリクルは繊維状弾性タンパク質で構成されていますが、セルロースでは構成されていません。 ペリクルは体の明確な形状を維持しますが、体の形状の一時的な変化を可能にするのに十分な柔軟性があり、これらの形状の変化はメタボディまたはユーグレノイドの動きとして話されています。

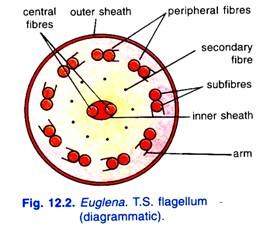
ペリクルの電子構造:
広告:
ペリクルの電子顕微鏡的研究は、それがらせん状に配置されたストリップで作られていることを明らかにする。 これらのストリップは、細胞体の両端で融合され、それぞれが一方の縁に沿って溝を有し、他方に沿って溝を有する。 隣接するストリップのエッジは、一方のストリップのリッジが他方のストリップの溝に収まるように重なり合って連結しています。
実際には、連結した尾根はペリクルの横紋の外観を与えます。 ストリップのすぐ下と平行に、粘液を分泌する粘液性の体の列と微小管の束が配置されていることが見出される(図10)。 12.3).

広告:
Cytostomeとcytopharynx:
前端には、中心の片側にわずかに漏斗状のcytostomeまたは細胞口があります。 Cytostomeは短い管状のcytopharynxまたはgulletにつながり、今度は大きな球形の小胞、貯水池または鞭毛嚢に合流する。 Cytostomeおよびcytopharynxは食糧の摂取のために貯蔵所からの液体の脱出のための運河として使用されません。
収縮性液胞:
広告:
大きな浸透圧調節体である収縮性液胞は、片側の貯水池の近くにある。 これは、おそらくより大きな液胞を形成するために一緒に融合し、いくつかの微細な付属収縮液胞に囲まれています。 収縮性液胞は、過剰の水および代謝のいくつかの老廃物をリザーバに排出し、そこから細胞小体を通って排出する。
鞭毛:
単一の長い鞭毛様鞭毛が細胞咽頭を通って細胞口から出てくる。 鞭毛の長さはミドリムシの種によって異なりますが、ミドリムシのviridisでは動物の体と同じくらい長いです。 これは、収縮性液胞の反対側から貯水池の基部から二つの根によって生じる。
各根は眼瞼形成細胞から湧出する(Gr.、眼瞼=まぶた; plastos=形成された)または細胞質の前部に埋め込まれた基底顆粒。
いくつかの作業者によると、二つの鞭毛があり、一つは長いものと短いものがあり、それぞれは貯水池の基部の細胞質に位置する基底顆粒から生じる。 短い鞭毛は貯水池の首を越えて伸びず、しばしば長い鞭毛に付着して分岐の外観を生じる。
鞭毛は外側の収縮性原形質鞘と内側の弾性軸フィラメントである軸索からなる。 鞭毛の遠位部分は、鞘の片側に沿って突出し、したがって、鞭毛はstichonematicタイプであるmastigonemesとして知られている多数の微細な繊維を含んでいます。
:
鞭毛の電子構造:
鞭毛の電子顕微鏡的研究により、それは2つの中央および9つの末梢線維からなることが明らかになった。 それぞれの中心線維は単一であり、末梢線維は対になっており、それぞれに二つの副線維を有する。 各末梢線維の2つのサブフィブリルの1つは、腕と呼ばれる短い突起の2列を持ち、すべての腕は同じ方向に向けられています。
二つの中心線維は内側の膜状鞘に囲まれているのが発見されている。 すべてのフィブリルは、細胞膜と連続した外側の原形質鞘内に囲まれている。 中心線維と末梢線維の間には九つの二次線維がある。
これらの線維はすべて融合して眼瞼形成細胞または基底顆粒に結合する。 Manton(1959)は、毛様収縮性繊維であるmastigonemesが、9つの末梢線維のうちの2つから生じることを示唆している。
汚名:
貯水池に近い細胞咽頭の内側の端付近には赤目斑または汚名があります。 それは脂質のしぶきの版、ヨウ素と青を汚すhaematochromeの赤い微粒としてカロテノイドの顔料から成っています。 柱頭はレンズとして作用する凹みの油性しぶきの無色の固まりとコップ型である。 柱頭は光に敏感です。
:
傍鞭毛体または光受容体:
傍鞭毛体として知られる小さな腫脹は、鞭毛の1つの根または2つの根の接合部にあります。 傍鞭毛体は光に敏感であり,光受容体と考えられている。 ChadefaudおよびProvasoliの最近の調査は柱頭およびparaflagellarボディが一緒に光受容器の器具を形作ることを示しました。
細胞質:
ユーグレナビリディスの細胞質は、外生生物の外層と内生生物の内層に分化している。 エクトプラズムは薄く、透明または非粒状であるが、内因性はより流動性で粒状である。 内生物は核、色素胞およびparamylum体を含む。
核:
ユーグレナは、単一の、大きな、円形または楕円形の小胞体の核を持ち、通常は体の中心付近または後端に向かって明確な位置にあります。 明確な核膜があります。 核はエンドソーム(核小体または核小体としても知られている)として知られている中心体を含んでいます。
クロマチンは核膜とエンドソームの間の空間に小さな顆粒を形成する。 大量の核生物が存在する。
色素体または葉緑体:
ユーグレナの体の中心から放射される、細長い色素体のようないくつかの、細長い、バンドがあります。 色素胞は緑の顔料、クロロフィルaを含み、βカロチンと共にbは、また葉緑体として知られています。
Euglena Viridisはこれらの色素胞から緑色を派生させています。 葉緑体は恒星のように配置されているか、星の光線のように配置されています。 各色素体または葉緑体は、ピレノイドによって囲まれているピレノフォアとして知られている非常に薄い中央部分からなる。
ピレノイドはパラミラムからなる一対の半球構造の間に囲まれている。 Paramylumはヨウ素との色を与えない多糖類(β-1、3グルカン)澱粉です。 葉緑体の慎重な観察は、それらの中にラメラまたはチラコイドを有する葉緑素の群の存在を示唆している。
各チラコイドは三つのラメラを持つ。; チラコイドは葉緑体の間質またはマトリックスに配置され、リボソームと脂肪球も含まれています。 葉緑体は三重膜エンベロープによって囲まれている。

パラミラム体:
様々な形や大きさのパラミラム体が内生生物全体に散在している。 これらは屈折体であり、光合成の産物であるparamylumの形で貯蔵された食品材料を含む。
その他:
細胞質には、ゴルジ装置、小胞体、ミトコンドリアのような他の細胞成分も含まれており、その数はリザーバーの近くにあり、リボソームは小胞体、小胞体、葉緑体に散在している。
4. 1869>
(i)鞭毛運動
(ii)ユーグレノイド運動
(i)鞭毛運動:
VickermanとCox(1967)は、鞭毛が移動に直接寄与していることを示唆しています。 しかし、鞭毛運動のメカニズムを説明するためにいくつかの理論が提唱されている。 Butschliは、鞭毛が一連の横方向の動きを受け、そうすることで、圧力がその表面に対して直角に水に作用することを観察した。
この圧力は、本体の主軸に対して、一方が平行に向けられ、他方が直角に向けられた二つの力を生成します。 平行力は動物を前方に駆動し、直角に作用する力は動物をそれ自身の軸上で回転させる。

Gray(1928)は、一連の波が鞭毛の一方の端から他方の端に通過することを示唆した。 これらの波は、動きの方向に1つ、体の主軸を持つ円形の方向に1つの2つのタイプの力を作り出します。 前者は前方に動物を駆動し、後者は動物を回転させるだろう。
かなり長い間、鞭毛は鞭毛運動中に前方に向けられていると一般的に推定されていましたが、現在では、鞭毛は有効な脳卒中ではまっすぐで膨隆し、回復
最近、Lowndes(1941-43)は鞭毛が移動中に後方に向けられていることを指摘している。 Lowndesによると、一連の螺旋波は、速度と振幅の増加に伴って毎秒約12で基部から後方に向けられた鞭毛の先端に連続的に通過する。
波は鞭毛に沿って螺旋状に進行し、ミドリムシの体は一秒に一回回転する。 したがって、その移動において、それは直線の周りの螺旋状の経路をたどり、前進する。 動きの率は毎分3つのmmである。
しかし、鞭毛の動きは、そのすべての線維の収縮に関連しています。 これらの小繊維の収縮のためのエネルギーはblepharoplastsのmitochondriaで形作られるATPsから得られます。
(ii)ユーグレノイド運動またはメタボリー:
ユーグレナは時々非常に独特の遅いうねりの動きを示します。 収縮および拡張の蠕動波が前方から後端まで全身を通過し、動物は前方に移動する。 体は最初に前端で短く、より広くなり、次に中央で、そして後端で後端になる。
このタイプの運動をユーグレノイド運動と呼び、ゆっくりとした限られた運動が起こる。 ユーグレノイドの動きは、細胞質の収縮またはペリクルの下の細胞質に存在するミオネームの収縮によってもたらされる。
5. ミドリムシの栄養:
ミドリムシの栄養モードは、混合栄養性である。、栄養物はholophyticかsaprophyticまたは両方のモードによって達成されます。
(i)好中球性栄養または独立栄養性栄養:
ユーグレナでは、栄養の主なモードは好中球性または植物様である。 食品は、植物のように光合成的に製造され、色素胞に存在する二酸化炭素、光およびクロロフィルの助けを借りて製造される。 クロロフィルは、太陽光の存在下で二酸化炭素を炭素と酸素に分解する。
酸素は自由に設定され、炭素は保持され、水の要素と組み合わされてparamylumのような炭水化物(多糖類)を形成する。 パラミラムは、ヨウ素溶液では青色にならないため、澱粉とは異なります。 ユーグレナでは、予備の食物は屈折性パラミラム体の形で貯蔵され、その数はよく供給されたユーグレナに豊富である。
(ii)腐生性または腐生性の栄養:
日光がない場合、ユーグレナは腐生性、浸透圧栄養性または腐生性として知られる別の栄養様式によってその食物を このモードでは、動物は、その一般的な体表面を通して、動物の環境中の腐敗物質から溶液中のいくつかの有機物質を吸収する。 それらは窒素の源のために硝酸塩の代りにアンモニウムの塩を、要求する。
ユーグレナは、完全な暗闇の中で葉緑素を失うと、サプロゾイド栄養に耐えることができます。 通常、暗闇の中で失われたクロロフィルは光の中で回復します。 しかし、E.gracilisのような形態では、変化は永久的であり、すなわち、一度失われたクロロフィルは回復されない。 腐生栄養はまた正常なholophytic栄養物を補うかもしれません。
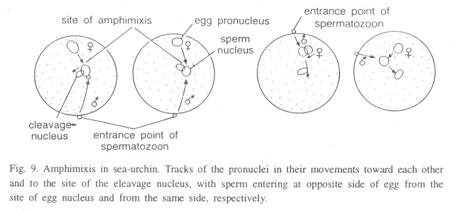
ピノサイトーシスは、タンパク質や他の大きな分子の摂取のために貯水池の基部でも発生することが報告されてい 生物が複数の方法を使用して展示する場合、それは栄養のmixotrophicモードを展示すると言われています。
ユーグレナはholophyticおよびsaprozoic栄養物を表わします、従って、栄養物のmixotrophicモードを表わします。 消化は、周囲の細胞質によって食物液胞に分泌される酵素によって行われる。
6. ユーグレナ-ヴィリディスの呼吸:
ユーグレナ-ビリディスでは、体表面を通る拡散によってガスの交換(O2の摂取とCO2の排出)が行われます。 周囲の水から溶存酸素を吸収し、拡散によって二酸化炭素を放出します。
日中、光合成中に放出された酸素は呼吸のために利用され、呼吸中に放出された二酸化炭素は光合成のために利用されると信じている理由はあ
7. ユーグレナ-ビリディスにおける排泄:
二酸化炭素と窒素性廃棄物(アンモニア)の除去は、一般的な体表面を介して拡散によって行われます。 しかし,少なくともいくらかの排泄は収縮性液胞によって行われる。
浸透圧調節:
ミドリムシは半透過性のペリクルを持ち、水中に生息しているため、内膜症によって水が体内に連続的に入る。 体内から過剰の水分を除去することは、浸透圧調節として知られています。 過剰の水分の除去は、収縮性液胞によって行われる。
副収縮液胞は周囲の細胞質から過剰な水分を収集し、その内容物を主収縮液胞に遊離させ、徐々にサイズが増加し、最終的に水を破裂させて貯水池に 貯水池の水から、細胞咽頭を通ってサイトソームによって脱出する。 これに伴い、水溶性廃棄物も体内から投げ出されます。
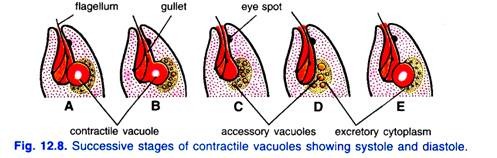
最近、Chadefaudは収縮液胞が特殊な顆粒状および排泄性細胞質に囲まれていることを指摘している。 収縮性液胞は周期的にその最大サイズに達し、その内容物を貯留層(すなわち収縮期)に排出するために崩壊する。
同時に、いくつかの小さな付属液胞が排泄細胞質に現れる。 これらの液胞はそれから新しい大きい液胞を形作るために一緒に溶けます(すなわち 最大サイズを達成し、前のもののように水を排出するために崩壊する拡張期)。
8. ミドリムシの行動:
ミドリムシは様々な刺激に応答し、非常にです。光に敏感です。 それは窓からのそれのような通常のライトの方に泳ぎ、強いライトを避ける。 ユーグレナの培養を調べると、ほとんどの動物が光に向かって側にあります。 光は、そのクロロフィルによる二酸化炭素の同化のために必要であるため、これは、動物にとって明確な利点である。
ミドリムシは直射日光を避けて泳ぎます。 長時間行動することが許されれば、直射日光は生物を殺すでしょう。 ミドリムシを含む皿を直射日光の下に置き、その半分を日陰にすると、動物は日陰の部分と直射日光を避け、それらに最も適した光の中で二つの間の小 12.9)、つまり、それらの最適です。
遊泳中のユーグレナは、自身の軸を中心に回転して回転する螺旋状に移動するが、光の方向が変わるたびに衝撃反応を示す。

目の前の領域は、体の他のどの部分よりも光に敏感であることがわかっています。 ユーグレナは、傍鞭毛体(光受容体)が柱頭またはeyespotによって陰影を付けられるたびに、光線に平行に配向する。 動物は、その位置を、光の方向に向かって、またはそこから離れて移動する方向に調整する。
動物が回転すると、柱頭はスクリーンとして機能し、横から光が当たると横鞭毛体が交互に露出または遮蔽されます。 動物は、傍鞭毛体が連続的に露出するまでそれ自身を調整するが、これは光源が前方または後方にまっすぐであるときに起こる。
ユーグレナは、試行錯誤パターン(phabotaxis)に機械的、熱的および化学的刺激に対する反応を回避する。 変化によって刺激されると、ユーグレナは、ほとんどの場合、停止または後方に移動し、背側表面に向かって強く回転するが、その長軸上で回転し続ける。
後端はピボットとして機能し、前端は水中の広い直径の円をトレースします。 動物は、この円の任意の点から新しい方向に前方に泳ぐことができます。 これは反応を避けています。
9. ミドリムシの繁殖:
ミドリムシは縦二分裂と多重分裂によって無性生殖を行う。 エンシストメントも行われます。 有性生殖は起こらないが、いくつかの種では原始的な形態が報告されている。
(i)縦二連核分裂:
活性期間中、水、温度および食物の入手可能性の好ましい条件下で、ユーグレナは縦方向の二成分核分裂によって再生する。 核分裂は常に対称的であり、すなわち、親のミドリムシは互いに正確に同一である二つの娘のミドリムシに分裂する。
核は有糸分裂によって分裂する。 エンドソームは横方向に伸長し、二つのほぼ等しい部分に収縮する。 核分裂は核膜内で起こる。
柱頭、眼瞼形成細胞、貯留層、細胞咽頭および色素胞およびパラミラム体などの前端のオルガネラも複製されている。 体は前端から後端まで縦に分裂し始め、その結果、二つの娘の個体が形成される。
古い鞭毛は半分に保持されますが、新しい鞭毛はもう一方によって発達し、収縮性液胞と傍鞭毛体は分裂しませんが、それらは消え、娘個体で再び作

(ii)複数の核分裂:
複数の核分裂は、通常、暗号化された状態で行われます。 時には、安静時または不活性期間中に、ユーグレナでエンシストが起こることがあります。 嚢胞内の細胞質および核の塊は、16または32の小さな娘個体を生じさせる繰り返し有糸分裂を受ける。
好条件が戻ると、嚢胞は壊れ、娘の個体は嚢胞から脱出する。 各娘の個体は様々な細胞小器官を発達させ、正常な生活を開始する。 いくつかの労働者は、娘の個体を胞子とみなし、このプロセスを胞子形成とみなした。
(iii)Palmella Stage:
時には、通常、不利な条件下では、多数のユーグレナが接近し、鞭毛を失い、丸くなることがあります。 それらは、それらが埋め込まれたままのゼラチン状の被覆または粘液マトリックスを分泌する。 この状態は、多くの場合、池の水面上の緑のスカムとして見られているパルメラステージと呼ばれています。
パルメラ段階の個体は代謝活動を続け、二元核分裂によって再現する。 好ましい条件の到着で、水のabsoprtionによってゼラチン状のカバーのうねりおよびeuglenaeは解放されます。 彼らは彼らの鞭毛を再生し、正常な活動的な生命を始める。
(iv)封じ込め:
干ばつ、極端な寒さ、極端な暑いなどの不利な条件の間、食物と酸素の不足ユーグレナは封じ込めを受ける。 まず第一に、ユーグレナは不活性になり、その鞭毛を失い、その周りに嚢胞を分泌する。 嚢胞は、ペリクルの下に横たわっている粘液体によって分泌される。
嚢胞は、ヘマトクロムと呼ばれる色素の存在のために、厚い壁、丸みを帯びた赤色である。 この嚢胞は保護タイプのものである。
エンシステッド条件の間、不利な条件の期間は正常に通過します。 エンシストメント中に、バイナリ核分裂は、嚢胞内の2-32小さな娘ユーグレナで、その結果、一つ以上の回を発生することがあります。 好ましい条件のリターンで、包嚢の壁の壊れ目は、動物活動的になり、包嚢から正常で自由な水泳の生命を導くために現れます。
実際には、エンシストメントは不利な条件の上に潮にのみ発生し、この条件の間にユーグレナの分散は広い領域に発生します。
10. ユーグレナビリディスの位置:
ユーグレナビリディスは、葉緑素やホロフィート栄養を持つ葉緑体などの植物の多くの特性を示しているが、以下の事実のために動物とみなされている:
(i)そのペリクルは植物のようにセルロースではなくタンパク質で作られている。
(ii)中心小体に匹敵する眼瞼形成細胞の存在。
(iii)柱頭および傍鞭毛体の存在、感光性構造。
(iv)植物には見られない収縮性液胞の存在。
(v)特定の動物学者によって主張されているように、栄養のSaprozoicモードとholozoic。
(vi)植物には見られない縦方向の二元核分裂の存在。
11. 他のいくつかのユーグレノイド鞭毛虫:
(i)Euglena Gracilis:
それは小さく、細長い、長さが約50ミクロンを測定する紡錘形です。 葉緑体は大きく、平らで、板状であり、数は約十である。 それぞれの葉緑体は、タンパク質性ピレノイドを有する。 ユーグレナ・ヴィリディスとは異なり、かつて暗闇の中で失われた葉緑体を取り戻すことはできません。 その細胞質には、葉緑体と関連して多くのパラミラム体が含まれています(Fig. 年12月14日)。

(ii)Euglena Spirogyra:
長さ約95ミクロン、幅約18ミクロンを測定する大型です。 その体は細長く、紡錘形であり、後方にその体は尾として引き出される。 ピレノイドのない多数の、小さい、円盤状の葉緑体があります。 それは細胞質のparamylumボディによって特徴付けられます(Fig. 年12月14日)。
(iii)Astasia Longa:
これは典型的なユーグレノイドの形態であり、一般的にはユーグレナグラシリスの漂白形態であると考えられている。 葉緑体、柱頭および傍鞭毛体は見出されない。 葉緑体が存在しないために浸透圧栄養によって栄養を示し、その細胞質には多くのパラミラム体が含まれています(Fig. 年12月14日-)は、日本の元av女優。
(iv)Paranema Trichophorum:
やや切り株を持つミドリムシの鞭毛虫です。 かなり大きい微生物にphagotrophyによってholozoically与えることを信じます。 その2つの鞭毛のうち、1つは運動性で長く、もう1つは尾を引き、体表面に付着していることが判明しています。 柱頭と傍鞭毛体は見つかっていない。 その細胞質には食物液胞と多くの小さなparamylum体が含まれています。
それは、その細胞咽頭に三毛虫と呼ばれる付属の棒状の装置が存在することを特徴としている(図18)。 年12月14日)。