WERBUNG:
In diesem Artikel werden wir über Euglena Viridis diskutieren:- 1. Habitat und Lebensraum von Euglena Viridis 2. Kultur von Euglena Viridis 3. Struktur 4. Fortbewegung 5. Ernährung 6. Atmung 7. Ausscheidung 8. Verhalten 9. Reproduktion 10. Position 11. Einige andere euglenoide Flagellaten.
Inhalt:
- Gewohnheit und Lebensraum von Euglena Green
- Kultur von Euglena Green
- Struktur von Euglena Green
- Fortbewegung von Euglena Green
- Ernährung von Euglena Green
- Atmung in Euglena Green
- Ausscheidung von Euglena Green
- Verhalten von Euglena Green
- Fortpflanzung in Euglena Green
- Lage von Euglena Green
- Einige andere euglenoide Flagellaten
- 1. Habitat und Lebensraum von Euglena Green:
- 2. Kultur von Euglena Viridis:
- 3. Struktur von Euglena Viridis:
- 4. Fortbewegung in Euglena Viridis:
- 5. Ernährung von Euglena Viridis:
- 6. Atmung bei Euglena Viridis:
- 7. Ausscheidung in Euglena Viridis:
- 8. Verhalten von Euglena Viridis:
- 9. Fortpflanzung bei Euglena Viridis:
- 10. Position von Euglena Viridis:
- 11. Einige andere euglenoide Flagellaten:
1. Habitat und Lebensraum von Euglena Green:
Euglena green (Gr. eu = wahr; glene = Augapfel oder Augenpupille; L., viridis = grün) ist ein gewöhnlicher, einsamer und frei lebender Süßwasserflagellat. Es kommt in Süßwasserbecken, Teichen, Gräben und langsam fließenden Bächen vor. Es kommt in Hülle und Fülle vor, wo es eine beträchtliche Menge an Vegetation gibt.
ANZEIGEN:
Teiche in den gepflegten Gärten mit verfallenden stickstoffhaltigen organischen Stoffen wie Zweigen, Blättern und Gesichtern von Tieren usw., sind gute Quelle dieses Organismus. Es lebt im Allgemeinen mit den anderen Arten der Gattung. Sie sind manchmal so zahlreich, dass sie dem Wasser eine deutliche grünliche Farbe verleihen oder manchmal einen grünen Schaumfilm auf der Oberfläche des Teichwassers bilden.
2. Kultur von Euglena Viridis:
Die Kultur von Euglena Viridis kann im Labor leicht mit der folgenden Methode hergestellt werden. Kochen Sie etwas Kuh- oder Pferdemist in angelehntem destilliertem Wasser und lassen Sie ihn zwei Tage abkühlen. Geben Sie dann etwas Unkraut aus einem Teich mit Euglenae in das Glas und stellen Sie das Glas in die Nähe des gut beleuchteten Fensters. In wenigen Tagen erscheinen Euglenae in dieser stickstoffhaltigen Infusion.
3. Struktur von Euglena Viridis:
Form:
WERBUNG:
Euglena viridis ist länglich und spindelförmig. Das vordere Ende ist stumpf, der mittlere Teil ist breiter, während das hintere Ende spitz ist.
Größe:
Euglena viridis ist an der dicksten Stelle des Körpers etwa 40-60 Mikrometer lang und 14-20 Mikrometer breit.
Pellikel:
Der Körper ist von einem dünnen, flexiblen, zähen und starken kutikulären Periplasten oder Pellikel bedeckt, das unter der Plasmamembran liegt. Es hat schräge, aber parallele Streifen, die rundum Myoneme genannt werden. Aber nach Chadefaud (1937) besteht das Pellikel aus einer äußeren dünnen Schicht Epikutikula und einer inneren dicken Schicht Kutikula.
Beide Pellikelschichten sind am ganzen Körper vorhanden, aber nur der Epikutikula endet in einem anterior platzierten Zytopharynx und Reservoir.
Das Pellikel besteht aus faserigem elastischem Protein, jedoch nicht aus Cellulose. Das Pellikel behält eine bestimmte Form des Körpers bei, ist aber flexibel genug, um vorübergehende Veränderungen der Körperform zuzulassen, diese Formänderungen werden als Metabody- oder Euglenoid-Bewegungen bezeichnet.

Elektronenstruktur des Pellets:
WERBUNG:
Die elektronenmikroskopische Untersuchung des Pellets zeigt, dass es aus spiralförmig angeordneten Streifen besteht. Diese Streifen sind an beiden Enden des Zellkörpers verschmolzen und haben jeweils eine Nut entlang einer Kante und eine Nut entlang der anderen. Die Kanten benachbarter Streifen überlappen sich und artikulieren sich so, dass der Grat eines Streifens in die Nut des anderen passt.
Tatsächlich verleihen die Gelenkkämme dem Pellikel ein gestreiftes Aussehen. Direkt unter und parallel zu den Streifen befindet sich eine Reihe von schleimabsondernden Schleimkörpern und Bündeln von Mikrotubuli (Abb. 12.3).

WERBUNG:
Zytostom und Zytopharynx:
Am vorderen Ende befindet sich ein trichterförmiges Zytostom oder ein Zellmund leicht seitlich des Zentrums. Das Zytostom führt in einen kurzen tubulären Zytopharynx oder eine Speiseröhre, die sich wiederum mit einem großen kugelförmigen Vesikel, dem Reservoir oder Flagellarsack, verbindet. Das Zytostom und der Zytopharynx dienen nicht zur Nahrungsaufnahme, sondern als Kanal zum Entweichen von Flüssigkeit aus dem Reservoir.
Kontraktile Vakuole:
WERBUNG:
Ein großer osmoregulatorischer Körper, die kontraktile Vakuole, liegt auf einer Seite in der Nähe des Reservoirs. Es ist von mehreren winzigen akzessorischen kontraktilen Vakuolen umgeben, die wahrscheinlich zu einer größeren Vakuole verschmelzen. Die kontraktile Vakuole entlädt den Überschuss an Wasser und einige Abfallprodukte des Stoffwechsels in das Reservoir, von wo aus es durch das Zytostom austritt.
Flagellum:
Ein einzelnes, langes, peitschenartiges Flagellum tritt durch den Zytopharynx aus dem Zytostom aus. Die Länge des Flagellums unterscheidet sich bei verschiedenen Euglena-Arten, bei Euglena viridis ist es jedoch so lang wie der Körper des Tieres. Es entsteht durch zwei Wurzeln von der Basis des Reservoirs von der der kontraktilen Vakuole gegenüberliegenden Seite.
Jede Wurzel entspringt einem Blepharoplast (Gr., blepharon = Augenlid; plastos = gebildet) oder Basalgranulat, das im vorderen Teil des Zytoplasmas eingebettet liegt.
Einigen Arbeitern zufolge gibt es zwei Flagellen, eine lange und eine kurze, die jeweils aus einem Basalgranulat stammen, das sich im Zytoplasma am Boden des Reservoirs befindet. Das kurze Flagellum erstreckt sich nicht über den Hals des Reservoirs hinaus und haftet häufig am langen Flagellum, wodurch das Auftreten einer Gabelung entsteht.
Das Flagellum besteht aus einer äußeren kontraktilen protoplasmatischen Hülle und einem inneren elastischen axialen Filament, dem Axonem. Der distale Teil des Flagellums enthält zahlreiche winzige Fasern, die als Mastigoneme bekannt sind und entlang einer Seite der Hülle vorstehen, und, deshalb, Das Flagellum ist stichonematisch.
ANZEIGEN:
Elektronenstruktur des Flagellums:
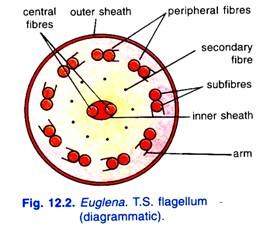
Die elektronenmikroskopische Untersuchung des Flagellums zeigt, dass es aus zwei zentralen und neun peripheren Fibrillen besteht. Jede zentrale Fibrille ist einzeln, während die peripheren Fibrillen mit jeweils zwei Subfibrillen gepaart sind. Eine der beiden Subfibrillen jeder peripheren Fibrille trägt eine doppelte Reihe von kurzen Vorsprüngen, die Arme genannt werden; Alle Arme sind in die gleiche Richtung gerichtet.
Die beiden zentralen Fibrillen befinden sich in einer inneren Membranhülle. Alle Fibrillen sind in einer äußeren protoplasmatischen Hülle eingeschlossen, die mit der Zellmembran durchgehend ist. Es gibt neun sekundäre Fibrillen zwischen zentralen und peripheren Fibrillen.
Alle diese Fibrillen verschmelzen zum Blepharoplasten oder Basalgranulat. Manton (1959) hat vorgeschlagen, dass Mastigoneme, haarartige kontraktile Fasern, aus zwei der neun peripheren Fibrillen entstehen.
Stigma:
In der Nähe des inneren Endes des Zytopharynx in der Nähe des Reservoirs befindet sich ein roter Augenfleck oder ein Stigma. Es besteht aus einer Platte aus Lipidtröpfchen, einem Carotinoidpigment als rotes Hämatochromgranulat, das mit Jod blau färbt. Stigma ist schalenförmig mit einer farblosen Masse öliger Tröpfchen in seiner Konkavität, die als Linse fungieren. Das Stigma ist lichtempfindlich.
ANZEIGEN:
Paraflagellarer Körper oder Photorezeptor:
Eine kleine Schwellung, die als paraflagellarer Körper bekannt ist, liegt entweder an einer Wurzel oder an der Verbindung zweier Wurzeln des Flagellums. Der paraflagellare Körper ist lichtempfindlich und gilt als Photorezeptor. Neuere Studien von Chadefaud und Provasoli haben gezeigt, dass das Stigma und der paraflagellare Körper zusammen den Photorezeptorapparat bilden.
Zytoplasma:
Das Zytoplasma von Euglena Viridis wird in eine äußere Schicht Ektoplasma und eine innere Schicht Endoplasma unterschieden. Das Ektoplasma ist dünn, klar oder nicht körnig, während das Endoplasma flüssiger und körniger ist. Das Endoplasma enthält Kern, Chromatophore und Paramylum-Körper.
Nucleus:
Euglena hat einen einzelnen, großen, runden oder ovalen und vesikulären Kern, der in einer bestimmten Position liegt, normalerweise nahe der Mitte oder zum hinteren Ende des Körpers hin. Es gibt eine ausgeprägte Kernmembran. Der Kern enthält einen zentralen Körper, der als Endosom bekannt ist (der auch als Nukleolus oder Karyosom bekannt ist).
Chromatin bildet kleine Körnchen im Raum zwischen Kernmembran und dem Endosom. Es gibt eine große Menge an Nukleoplasma.
Chromatophore oder Chloroplasten:
Von der Körpermitte der Euglena strahlen mehrere, schlanke, bandförmige, längliche Chromatophore aus. Die Chromatophore enthalten das grüne Pigment, Chlorophyll a und b, zusammen mit β-Carotin und werden auch als Chloroplasten bezeichnet.
Euglena Viridis leitet ihre grüne Farbe von diesen Chromatophoren ab. Chloroplasten sind stellar angeordnet oder wie die Strahlen der Sterne. Jeder Chromatophor oder Chloroplast besteht aus einem sehr dünnen zentralen Teil, der als Pyrenophor bekannt ist und von einem Pyrenoid eingeschlossen ist.
Das Pyrenoid ist zwischen einem Paar halbkugelförmiger Strukturen aus Paramylum eingeschlossen. Paramylum ist eine Polysaccharidstärke (β-1,3-Glucan), die mit Jod keine Farbe ergibt. Eine sorgfältige Beobachtung von Chloroplasten deutet auf das Vorhandensein von Gruppen von chlorophylltragenden Lamellen oder Thylakoiden in ihnen hin.
Jedes Thylakoid trägt drei Lamellen; die Thylakoide befinden sich im Stroma oder in der Matrix der Chloroplasten und enthalten auch Ribosomen und Fettkügelchen. Ein Chloroplast wird durch eine dreifache Membranhülle begrenzt.

Paramylum-Körper:
Paramylum-Körper verschiedener Formen und Größen werden im gesamten Endoplasma verstreut gefunden. Diese sind refraktile Körper und enthalten gespeichertes Nahrungsmittelmaterial in Form von Paramylum, das ein Produkt der Photosynthese ist.
Andere zytoplasmatische Gehalte:
Das Zytoplasma enthält auch andere zelluläre Komponenten wie Golgi-Apparate, endoplasmatisches Retikulum, Mitochondrien, deren Anzahl näher am Reservoir liegt, und die Ribosomen, die verstreut im Endoplasma, am endoplasmatischen Retikulum und in den Chloroplasten gefunden werden.
4. Fortbewegung in Euglena Viridis:
Es gibt zwei Methoden der Fortbewegung in Euglena Viridis, nämlich:
(i) Flagellar Bewegung
(ii) Euglenoid Bewegung
(i) Flagellar Bewegung:
Vickerman und Cox (1967) haben vorgeschlagen, dass das Flagellum einen direkten Beitrag zur Fortbewegung leistet. Es wurden jedoch mehrere Theorien aufgestellt, um den Mechanismus der Flagellenbewegung zu erklären. Butschli beobachtete, dass das Flagellum eine Reihe von seitlichen Bewegungen durchläuft und dabei ein Druck auf das Wasser im rechten Winkel zu seiner Oberfläche ausgeübt wird.
Dieser Druck erzeugt zwei Kräfte, die eine parallel und die andere rechtwinklig zur Hauptachse des Körpers gerichtet sind. Die parallele Kraft treibt das Tier vorwärts und die im rechten Winkel wirkende Kraft würde das Tier um seine eigene Achse drehen.

Gray (1928) schlug vor, dass eine Reihe von Wellen von einem Ende des Flagellums zum anderen verläuft. Diese Wellen erzeugen zwei Arten von Kräften, eine in Bewegungsrichtung und die andere in Kreisrichtung mit der Hauptachse des Körpers. Ersteres treibt das Tier vorwärts und letzteres würde das Tier drehen.
Lange Zeit wurde allgemein angenommen, dass das Flagellum während der Flagellenbewegung nach vorne gerichtet ist, aber jetzt ist man sich allgemein einig, dass das Flagellum im effektiven Schlag gerade und prall ist und im Erholungsschlag nach hinten fällt.
Kürzlich hat Lowndes (1941-43) darauf hingewiesen, dass das Flagellum während der Fortbewegung nach hinten gerichtet ist. Nach Lowndes verläuft eine Reihe von Spiralwellen nacheinander von der Basis zur Spitze des nach hinten gerichteten Flagellums mit etwa 12 pro Sekunde mit zunehmender Geschwindigkeit und Amplitude.
Die Wellen verlaufen spiralförmig entlang des Flagellums und bewirken, dass sich der Körper von Euglena einmal in einer Sekunde dreht. So verfolgt es in seiner Fortbewegung einen spiralförmigen Pfad um eine gerade Linie und bewegt sich vorwärts. Die Bewegungsrate beträgt 3 mm pro Minute.
Die Bewegung des Flagellums hängt jedoch mit der Kontraktion aller Fibrillen zusammen. Die Energie für die Kontraktion dieser Fibrillen stammt von ATPs, die in den Mitochondrien von Blepharoplasten gebildet werden.
(ii) Euglenoide Bewegung oder Metaboly:
Euglena zeigt manchmal eine sehr eigenartige langsame zappelnde Bewegungen. Eine peristaltische Welle der Kontraktion und Expansion verläuft vom vorderen zum hinteren Ende über den gesamten Körper und das Tier bewegt sich vorwärts. Der Körper wird zuerst am vorderen Ende kürzer und breiter, dann in der Mitte und später am hinteren Ende.
Diese Art von Bewegung wird als euglenoide Bewegung bezeichnet, bei der langsame und begrenzte Bewegungen auftreten. Euglenoide Bewegungen werden entweder durch die Kontraktionen des Zytoplasmas oder durch die Kontraktionen von Myonemen hervorgerufen, die im Zytoplasma unterhalb des Pellikels vorhanden sind.
5. Ernährung von Euglena Viridis:
Die Art der Ernährung in Euglena ist mixotroph, d.h. Wird die Ernährung entweder durch holophytische oder saprophytische oder durch beide Modi erreicht.
(i) Holophytische oder autotrophe Ernährung:
Bei Euglena ist die Hauptnahrung holophytisch oder pflanzenähnlich. Die Nahrung wird wie in Pflanzen photosynthetisch mit Hilfe von Kohlendioxid, Licht und Chlorophyll in den Chromatophoren hergestellt. Das Chlorophyll zersetzt das Kohlendioxid in Gegenwart von Sonnenlicht in Kohlenstoff und Sauerstoff.
Der Sauerstoff wird freigesetzt und Kohlenstoff wird zurückgehalten und mit den Elementen des Wassers kombiniert, um Kohlenhydrat (Polysaccharid) wie Paramylum zu bilden. Das Paramylum unterscheidet sich von Stärke, weil es mit Jodlösung nicht blau wird. In Euglena wird das Reservefutter in Form von refraktiven Paramylum-Körpern gelagert und ihre Anzahl ist in einer gut gefütterten Euglena reichlich vorhanden.
(ii) Saprophytische oder saprozoische Ernährung:
In Abwesenheit von Sonnenlicht bezieht Euglena seine Nahrung durch eine andere Art der Ernährung, die als saprophytisch, osmotroph oder saprozoisch bekannt ist. In diesem Modus absorbiert das Tier durch seine allgemeine Körperoberfläche einige organische Substanzen in Lösung von zerfallenden Stoffen in der Umgebung des Tieres. Sie benötigen Ammoniumsalze anstelle von Nitraten für ihre Stickstoffquellen.
Euglena kann von saprozoischer Nahrung leben, wenn sie in völliger Dunkelheit ihr Chlorophyll verliert. Normalerweise werden die Chlorophylle, die in der Dunkelheit verloren gehen, im Licht wiedergewonnen. Aber in Formen wie E. gracilis ist die Veränderung dauerhaft, d. H. Die Chlorophylle, die einmal verloren gegangen sind, werden nicht wiedergewonnen. Die saprophytische Ernährung kann auch die normale holophytische Ernährung ergänzen.
Es wurde auch berichtet, dass eine Pinozytose an der Basis des Reservoirs für die Aufnahme von Proteinen und anderen großen Molekülen auftritt. Wenn ein Organismus mit mehr als einer Methode exponiert, dann wird gesagt, dass er eine mixotrophe Ernährungsweise aufweist.
Euglena zeigt sowohl holophytische als auch saprozoische Ernährung, daher zeigt es eine mixotrophe Ernährungsweise. Die Verdauung erfolgt durch Enzyme, die vom umgebenden Zytoplasma in die Nahrungsvakuolen abgesondert werden.
6. Atmung bei Euglena Viridis:
Bei Euglena Viridis erfolgt der Gasaustausch (Aufnahme von O2 und Abgabe von CO2) durch Diffusion durch die Körperoberfläche. Es absorbiert gelösten Sauerstoff aus dem umgebenden Wasser und gibt Kohlendioxid durch Diffusion ab.
Es gibt allen Grund zu der Annahme, dass während der Tageszeit der bei der Photosynthese freigesetzte Sauerstoff zur Atmung und das bei der Atmung abgegebene Kohlendioxid zur Photosynthese genutzt werden kann.
7. Ausscheidung in Euglena Viridis:
Die Ausscheidung von Kohlendioxid und stickstoffhaltigem Abfallprodukt (Ammoniak) erfolgt durch Diffusion durch die allgemeine Körperoberfläche. Zumindest eine gewisse Ausscheidung erfolgt jedoch durch die kontraktile Vakuole.
Osmoregulation:
Da Euglena Viridis ein semipermeables Pellikel hat und im Wasser lebt, gelangt Wasser kontinuierlich durch Endosmose in seinen Körper. Die Entfernung von überschüssigem Wasser aus dem Körper wird als Osmoregulation bezeichnet. Die Beseitigung von überschüssigem Wasser erfolgt durch die kontraktile Vakuole.
Die akzessorischen kontraktilen Vakuolen sammeln überschüssiges Wasser aus dem umgebenden Zytoplasma und setzen ihren Inhalt in die kontraktile Hauptvakuole frei, die allmählich an Größe zunimmt und schließlich platzt und das Wasser in das Reservoir drückt. Aus dem Reservoir entweicht Wasser durch Zytosomen durch den Zytopharynx. Gleichzeitig werden auch wasserlösliche Abfälle aus dem Körper geworfen.
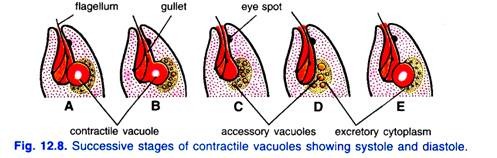
Kürzlich hat Chadefaud darauf hingewiesen, dass die kontraktile Vakuole von einem spezialisierten körnigen und ausscheidenden Zytoplasma umgeben ist. Die kontraktile Vakuole erreicht periodisch ihre maximale Größe und kollabiert, um ihren Inhalt in das Reservoir abzugeben (d. H. Systole).
Gleichzeitig erscheinen mehrere kleine akzessorische Vakuolen im Ausscheidungszytoplasma. Diese Vakuolen verschmelzen dann zu einer neuen großen Vakuole (d.h., Diastole), die die maximale Größe erreicht und zusammenbricht, um das Wasser wie das erstere abzulassen.
8. Verhalten von Euglena Viridis:
Euglena Viridis reagiert auf eine Vielzahl von Reizen und ist sehr empfindlich .empfindlich gegen Licht. Es schwimmt auf ein gewöhnliches Licht wie das eines Fensters zu und vermeidet starkes Licht. Wenn eine Kultur von Euglena untersucht wird, werden die meisten Tiere auf der Seite zum Licht gefunden. Dies ist für das Tier von deutlichem Vorteil, da Licht für die Aufnahme von Kohlendioxid durch sein Chlorophyll notwendig ist.
Euglena schwimmt weg von den direkten Sonnenstrahlen. Direkte Sonneneinstrahlung tötet den Organismus ab, wenn er längere Zeit einwirken kann. Wenn eine Schale, die Euglenae enthält, in das direkte Sonnenlicht gestellt wird und dann eine Hälfte davon beschattet wird, meiden die Tiere den schattigen Teil und auch das direkte Sonnenlicht und bleiben in einem kleinen Band zwischen den beiden in dem für sie am besten geeigneten Licht (Abb. 12.9), das heißt, ihr Optimum.
Eine schwimmende Euglena bewegt sich spiralförmig um ihre eigene Achse, zeigt jedoch eine Schockreaktion, wenn die Lichtrichtung geändert wird.

Es wurde festgestellt, dass die Region vor dem Augenfleck lichtempfindlicher ist als jeder andere Körperteil. Euglena orientiert sich parallel zu den Lichtstrahlen, wenn der paraflagellare Körper (Photorezeptor) durch das Stigma oder den Augenfleck beschattet wird. Das Tier passt seine Position an die Richtung des Lichts an, das sich entweder auf es zu oder von ihm weg bewegt.
Wenn sich das Tier dreht, wirkt das Stigma als Schirm, der paraflagellare Körper wird abwechselnd freigelegt oder abgeschirmt, wenn Licht von der Seite darauf fällt. Das Tier passt sich an, bis der paraflagellare Körper kontinuierlich exponiert ist, dies geschieht, wenn die Lichtquelle entweder direkt vor oder hinter ist.
Euglena reagiert auf mechanische, thermische und chemische Reize nach einem Trial-and-Error-Muster (Phabotaxis). Wenn Euglena durch eine Veränderung stimuliert wird, stoppt oder bewegt sie sich in den meisten Fällen rückwärts, dreht sich stark in Richtung der dorsalen Oberfläche, dreht sich aber weiterhin um ihre lange Achse.
Das hintere Ende fungiert dann als Drehpunkt, während das vordere Ende einen Kreis mit großem Durchmesser im Wasser nachzeichnet. Das Tier kann von jedem Punkt in diesem Kreis in eine neue Richtung vorwärts schwimmen. Dies vermeidet Reaktionen.
9. Fortpflanzung bei Euglena Viridis:
Euglena Viridis vermehrt sich asexuell durch longitudinale binäre Spaltung und multiple Spaltung. Encystment findet ebenfalls statt. Sexuelle Fortpflanzung tritt nicht auf, obwohl eine primitive Form davon bei einigen Arten berichtet wird.
(i) Longitudinale binäre Spaltung:
Während aktiver Perioden vermehrt sich Euglena unter günstigen Bedingungen von Wasser, Temperatur und Nahrungsverfügbarkeit durch longitudinale binäre Spaltung. Die Spaltung ist immer symmetrogen, d.h. die Eltern-Euglena teilt sich in zwei Tochter-Euglenae, die genau identisch sind.
Der Kern teilt sich durch Mitose. Das Endosom verlängert sich quer und wird in zwei ungefähr gleiche Teile verengt. Die Kernteilung findet innerhalb der Kernmembran statt.
Die Organellen am vorderen Ende wie Stigma, Blepharoplasten, Reservoir, Zytopharynx und Chromatophore und Paramylum-Körper sind ebenfalls dupliziert. Der Körper beginnt sich der Länge nach vom vorderen Ende nach unten zum hinteren Ende zu teilen, was zur Bildung von zwei Tochterpersonen führt.
Das alte Flagellum wird von der einen Hälfte beibehalten, während ein neues Flagellum von der anderen entwickelt wird, kontraktile Vakuole und paraflagellarer Körper teilen sich nicht, sondern verschwinden und werden bei den Tochterpersonen wieder hergestellt.

( ii) Mehrfachspaltung:
Die Mehrfachspaltung findet normalerweise im verschlüsselten Zustand statt. Manchmal tritt in Ruhe- oder inaktiven Perioden eine Einkreisung in Euglena auf. Die Masse des Zytoplasmas und der Kern innerhalb der Zyste werden wiederholt mitotisch geteilt, wodurch 16 oder 32 kleine Tochterpersonen entstehen.
Bei Rückkehr günstiger Bedingungen bricht die Zyste und die Tochterpersonen entweichen aus der Zyste. Jede Tochter Individuum entwickelt die verschiedenen Organellen und beginnt das normale Leben. Einige Arbeiter betrachteten die Tochterpersonen als Sporen und diesen Prozess als Sporulation.
(iii) Palmella-Stadium:
Manchmal, meist unter ungünstigen Bedingungen, kommt eine große Anzahl von Euglenae nahe zusammen, verliert ihre Flagellen und wird gerundet. Sie scheiden eine gallertartige Hülle oder eine schleimige Matrix aus, in die sie eingebettet bleiben. Dieser Zustand wird als Palmella-Stadium bezeichnet, das oft als grüner Abschaum auf der Wasseroberfläche von Teichen angesehen wird.
Individuen des Palmella-Stadiums führen metabolische Aktivitäten durch und vermehren sich durch binäre Spaltung. Bei günstigen Bedingungen schwillt die gallertartige Hülle durch die Absopration von Wasser an und die Euglenae werden freigesetzt. Sie regenerieren ihre Flagellen und beginnen ein normales aktives Leben.
( iv) Encystment:
Bei ungünstigen Bedingungen wie Dürre, extremer Kälte oder extremer Hitze, Nahrungsknappheit und Sauerstoffmangel wird Euglena encystment unterzogen. Zunächst wird Euglena inaktiv, verliert ihr Flagellum und scheidet eine Zyste aus. Die Zyste wird von den unter dem Pellikel liegenden Schleimkörpern abgesondert.
Die Zyste ist dickwandig, abgerundet und rot gefärbt, da ein Pigment namens Hämatochrome vorhanden ist. Diese Zyste ist vom schützenden Typ.
Während des verschlüsselten Zustands werden die Perioden ungünstiger Bedingungen erfolgreich bestanden. Während der encystment kann binäre Spaltung ein oder mehrere Male auftreten, was zu 2 bis 32 kleine Tochter euglenae innerhalb der Zyste. Bei Rückkehr günstiger Bedingungen bricht die Zystenwand, die Tiere werden aktiv und treten aus der Zyste aus, um ein normales freies Schwimmleben zu führen.
In der Tat tritt die Einkreisung nur auf, um die ungünstigen Bedingungen zu überbrücken, und während dieser Bedingung tritt eine Ausbreitung von Euglena auf ein weites Gebiet auf.
10. Position von Euglena Viridis:
Euglena Viridis zeigt viele Merkmale von Pflanzen wie Chloroplasten mit Chlorophyll und holophytischer Ernährung, wird jedoch aufgrund der folgenden Tatsachen als Tier angesehen:
(i) Sein Pellikel besteht aus Proteinen und nicht wie bei Pflanzen aus Cellulose.
(ii) Vorhandensein von Blepharoplasten, vergleichbar mit Zentriolen.
(iii) Vorhandensein von Stigma und Paraflagellarkörper, die lichtempfindlichen Strukturen.
(iv) Vorhandensein von kontraktilen Vakuolen, die in Pflanzen nicht vorkommen.
(v) Saprozoische Art der Ernährung und auch Holozoikum, wie von bestimmten Zoologen behauptet wurde.
(vi) Vorhandensein einer longitudinalen binären Spaltung, die in Pflanzen nicht vorkommt.
11. Einige andere euglenoide Flagellaten:
(i) Euglena Gracilis:
Es ist klein, länglich, spindelförmig und etwa 50 Mikrometer lang. Die Chloroplasten sind groß, flach, plattenförmig und etwa zehn an der Zahl. Jeder Chloroplast trägt ein Proteinpyrenoid. Im Gegensatz zu Euglena viridis können seine Chloroplasten, die einmal in der Dunkelheit verloren gegangen sind, nicht wiedergewonnen werden. Sein Zytoplasma enthält viele Paramyllkörper in Verbindung mit den Chloroplasten (Abb. 12.14 A).

( ii) Euglena Spirogyra:
Es ist großformatig und misst etwa 95 Mikrometer Länge und 18 Mikrometer Breite. Sein Körper ist länglich, spindelförmig und posterior ist sein Körper als Schwanz ausgezogen. Es gibt zahlreiche, kleine, scheibenartige Chloroplasten ohne Pyrenoide. Es zeichnet sich durch die Paramylum-Körper in seinem Zytoplasma aus (Abb. 12.14 B).
(iii) Astasia Longa:
Es ist eine typische euglenoide Form und soll allgemein die gebleichte Form von Euglena gracilis sein. Chloroplasten, Stigma und paraflagellarer Körper werden nicht gefunden. Es zeigt eine Ernährung durch Osmotrophie aufgrund der Abwesenheit von Chloroplasten und sein Zytoplasma enthält viele parametrische Körper (Abb. 12.14 C).
(iv) Paranema Trichophorum:
Es ist ein euglenoides Flagellat mit etwas stumpfem Körper. Es wird angenommen, dass es sich holozoisch durch Phagotrophie von ziemlich großen Mikroorganismen ernährt. Von seinen zwei Flagellen ist eine lokomotorisch und lang, während die andere nachläuft und an ihrer Körperoberfläche befestigt ist. Das Stigma und der paraflagellare Körper werden nicht gefunden. Sein Zytoplasma enthält Nahrungsvakuolen und viele kleine Paramylum-Körper.
Es ist gekennzeichnet durch das Vorhandensein eines stäbchenförmigen Zusatzapparates, der Trichite genannt wird, in seinem Zytopharynx (Abb. 12.14 D).