ANNONCES:
Dans cet article, nous discuterons d’Euglena Viridis: – 1. Port et habitat d’Euglena Viridis 2. Culture d’Euglena Viridis 3. Structure 4. Locomotion 5. Nutrition 6. Respiration 7. Excrétion 8. Comportement 9. Reproduction 10. Position 11. Quelques Autres Flagellés Euglénoïdes.
Contenu:
- Habitat et Habitat du Vert d’Euglène
- Culture du Vert d’Euglène
- Structure du Vert d’Euglène
- Locomotion du Vert d’Euglène
- Nutrition du Vert d’Euglène
- Respiration du Vert d’Euglène
- Excrétion du Vert d’Euglène
- Comportement du Vert d’Euglène
- Reproduction dans le Vert d’Euglène
- Position du Vert d’Euglène
- Quelques Autres Flagellés Euglénoïdes
- 1. Port et habitat du Vert d’Euglène :
- 2. Culture d’Euglena Viridis:
- 3. Structure d’Euglena Viridis:
- 4. Locomotion chez Euglena Viridis:
- 5. Nutrition d’Euglena Viridis:
- 6. Respiration à Euglena Viridis:
- 7. Excrétion chez Euglena Viridis:
- 8. Comportement d’Euglena Viridis:
- 9. Reproduction chez Euglena Viridis :
- 10. Position d’Euglena Viridis:
- 11. Quelques autres Flagellés Euglénoïdes :
1. Port et habitat du Vert d’Euglène :
Vert d’Euglène (Gr. eu = vrai; glene = boule oculaire ou pupille oculaire; L., viridis = vert) est un flagellé d’eau douce commun, solitaire et libre. On le trouve dans les mares d’eau douce, les étangs, les fossés et les ruisseaux qui coulent lentement. On le trouve en abondance là où il y a une quantité considérable de végétation.
ANNONCES:
Étangs dans les jardins bien entretenus contenant des matières organiques azotées en décomposition, telles que des brindilles, des feuilles et des visages d’animaux, etc., sont une bonne source de cet organisme. Il vit généralement avec les autres espèces du genre. Ils sont parfois si nombreux qu’ils donnent une couleur verdâtre distincte à l’eau ou forment parfois un film vert d’écume à la surface de l’eau de l’étang.
2. Culture d’Euglena Viridis:
La culture d’Euglena Viridis peut être facilement préparée en laboratoire par la méthode suivante. Faites bouillir de la bouse de vache ou de cheval dans de l’eau distillée à entrouverte et laissez-la refroidir pendant deux jours. Ensuite, mettez des mauvaises herbes d’un étang contenant des Euglènes dans le pot et placez le pot près de la fenêtre bien éclairée. Dans quelques jours, des Euglènes apparaîtront dans cette infusion azotée.
3. Structure d’Euglena Viridis:
Forme:
ANNONCES:
Euglena viridis est allongée et en forme de fuseau. L’extrémité antérieure est émoussée, la partie médiane est plus large, tandis que l’extrémité postérieure est pointue.
Taille:
Euglena viridis mesure environ 40 à 60 microns de longueur et 14 à 20 microns de largeur à la partie la plus épaisse du corps.
Pellicule:
Le corps est recouvert d’un périplaste ou d’une pellicule cuticulaire mince, flexible, dur et solide qui se trouve sous la membrane plasmique. Il a des stries obliques mais parallèles appelées myonèmes tout autour. Mais selon Chadefaud (1937), la pellicule est constituée d’une épicuticule à couche mince externe et d’une cuticule à couche épaisse interne.
Les deux couches de pellicule sont présentes sur tout le corps, mais seule l’épicuticule se termine par un cytopharynx et un réservoir placés antérieurement.
La pellicule est composée de protéines élastiques fibreuses mais pas de cellulose. La pellicule maintient une forme définie du corps, mais elle est suffisamment flexible pour permettre des changements temporaires de la forme du corps, ces changements de forme sont appelés mouvements métabody ou euglénoïdes.

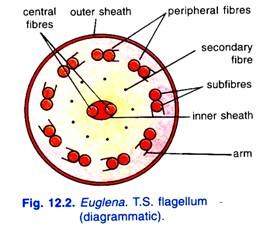
Structure électronique de la pellicule:
ANNONCES:
L’étude microscopique électronique de la pellicule révèle qu’elle est constituée de bandes disposées en hélice. Ces bandes sont fusionnées aux deux extrémités du corps de cellule et présentent chacune une rainure le long d’un bord et une rainure le long de l’autre. Les bords des bandes voisines se chevauchent et s’articulent de manière à ce que l’arête d’une bande s’insère dans la rainure de l’autre.
En effet, les arêtes articulées donnent à la pellicule un aspect strié. Juste en dessous et parallèlement aux bandes, une rangée de corps mucifères sécréteurs de mucus et des faisceaux de microtubles sont disposés (Fig. 12.3).

ANNONCES:
Cytostome et cytopharynx:
À l’extrémité antérieure se trouve un cytostome ou une bouche cellulaire en forme d’entonnoir légèrement sur un côté du centre. Le cytostome conduit à un court cytopharynx tubulaire ou à un œsophage qui, à son tour, rejoint une grande vésicule sphérique, le réservoir ou le sac flagellaire. Le cytostome et le cytopharynx ne sont pas utilisés pour l’ingestion de nourriture mais comme canal d’évacuation du liquide du réservoir.
Vacuole contractile:
ANNONCES:
Un grand corps osmorégulateur, la vacuole contractile se trouve près du réservoir d’un côté. Il est entouré de plusieurs minuscules vacuoles contractiles accessoires, qui fusionnent probablement ensemble pour former la plus grande vacuole. La vacuole contractile rejette l’excès d’eau et certains déchets du métabolisme dans le réservoir d’où il sort par le cytostome.
Flagelle:
Un seul flagelle long en forme de fouet émerge du cytostome par le cytopharynx. La longueur du flagelle diffère selon les espèces d’Euglena, mais chez Euglena viridis, elle est aussi longue que le corps de l’animalcule. Il provient de deux racines de la base du réservoir du côté opposé à la vacuole contractile.
Chaque racine jaillit d’un blépharoplaste (Gr., blépharon = paupière; plastos = formé) ou granule basal qui se trouve noyé dans la partie antérieure du cytoplasme.
Selon certains travailleurs, il existe deux flagelles, l’un long et l’autre court, provenant chacun d’un granule basal situé dans le cytoplasme à la base du réservoir. Le flagelle court ne s’étend pas au-delà du col du réservoir et il adhère souvent au flagelle long produisant l’apparence d’une bifurcation.
Le flagelle est constitué d’une gaine protoplasmique contractile externe et d’un filament axial élastique interne, l’axonème. La partie distale du flagelle contient de nombreuses fibres minuscules appelées mastigonèmes qui font saillie le long d’un côté de la gaine et, par conséquent, le flagelle est de type stichonématique.
ANNONCES:
Structure électronique du flagelle:
L’étude au microscope électronique du flagelle révèle qu’il se compose de deux fibrilles centrales et de neuf fibrilles périphériques. Chaque fibrille centrale est unique, tandis que les fibrilles périphériques sont appariées avec deux sous-fibrilles dans chacune. L’une des deux sous-fibrilles de chaque fibrille périphérique porte une double rangée de courtes saillies appelées bras ; tous les bras étant dirigés dans le même sens.
Les deux fibrilles centrales se trouvent enfermées dans une gaine membraneuse interne. Toutes les fibrilles sont enfermées dans une gaine protoplasmique externe continue avec la membrane cellulaire. Il y a neuf fibrilles secondaires entre les fibrilles centrales et périphériques.
Toutes ces fibrilles fusionnent pour rejoindre le blépharoplaste ou le granule basal. Manton (1959) a suggéré que les mastigonèmes, des fibres contractiles ressemblant à des poils, proviennent de deux des neuf fibrilles périphériques.
Stigmate:
Près de l’extrémité interne du cytopharynx, près du réservoir, se trouve une tache ou un stigmate des yeux rouges. Il se compose d’une plaque de gouttelettes lipidiques, un pigment caroténoïde sous forme de granules rouges d’hématochrome qui se colore en bleu avec de l’iode. Le stigmate est en forme de coupe avec une masse incolore de gouttelettes huileuses dans sa concavité qui fonctionnent comme une lentille. Le stigmate est sensible à la lumière.
ANNONCES:
Corps paraflagellaire ou photorécepteur:
Un petit gonflement connu sous le nom de corps paraflagellaire se trouve soit sur une racine, soit à la jonction de deux racines du flagelle. Le corps paraflagellaire est sensible à la lumière et il est considéré comme photorécepteur. Des études récentes sur Chadefaud et Provasoli ont montré que le stigmate et le corps paraflagellaire forment ensemble l’appareil photorécepteur.
Cytoplasme:
Le cytoplasme d’Euglena Viridis est différencié en une couche externe d’ectoplasme et une couche interne d’endoplasme. L’ectoplasme est mince, clair ou non granulaire, tandis que l’endoplasme est plus fluide et granulaire. L’endoplasme contient des corps de noyau, de chromatophores et de paramyle.
Noyau:
L’Euglène a un noyau unique, grand, rond ou ovale et vésiculaire situé dans une position définie généralement près du centre ou vers l’extrémité postérieure du corps. Il existe une membrane nucléaire distincte. Le noyau contient un corps central appelé endosome (également appelé nucléole ou caryosome).
La chromatine forme de petits granules dans l’espace entre la membrane nucléaire et l’endosome. Il y a une grande quantité de nucléoplasme.
Chromatophores ou chloroplastes:
Rayonnant du centre du corps d’Euglena, il y a plusieurs chromatophores allongés, minces, en forme de bande. Les chromatophores contiennent le pigment vert, la chlorophylle a et b, ainsi que le β-carotèneet sont également connus sous le nom de chloroplastes.
Euglena Viridis tire sa couleur verte de ces chromatophores. Les chloroplastes sont disposés de manière stellaire ou comme les rayons des étoiles. Chaque chromatophore ou chloroplaste est constitué d’une partie centrale très mince appelée pyrénophore qui est entourée d’un pyrénoïde.
Le pyrénoïde est enfermé entre une paire de structures hémisphériques en paramyle. Le paramylum est un amidon polysaccharidique (β-1,3 glucane) qui ne donne pas de couleur avec l’iode. Une observation attentive des chloroplastes suggère la présence de groupes de chlorophylle portant des lamelles ou des thylakoïdes.
Chaque thylakoïde porte trois lamelles; les thylakoïdes sont placés dans le stroma ou la matrice des chloroplastes et contiennent également des ribosomes et des globules gras. Un chloroplaste est délimité par une triple enveloppe membranaire.

Corps de paramyle:
Des corps de paramyle de différentes formes et tailles se trouvent dispersés dans tout l’endoplasme. Ce sont des corps réfractiles et contiennent du matériel alimentaire stocké sous forme de paramyle qui est un produit de la photosynthèse.
Autres contenus cytoplasmiques:
Le cytoplasme contient également d’autres composants cellulaires comme les appareils de Golgi, le réticulum endoplasmique, les mitochondries dont le nombre est plus proche du réservoir et les ribosomes qui se trouvent dispersés dans l’endoplasme, sur le réticulum endoplasmique et dans les chloroplastes.
4. Locomotion chez Euglena Viridis:
Il existe deux méthodes de locomotion chez Euglena Viridis, à savoir:
(i) Mouvement flagellaire
(ii) Mouvement euglénoïde
(i) Mouvement flagellaire:
Vickerman et Cox (1967) ont suggéré que le flagelle contribue directement à la locomotion. Cependant, plusieurs théories ont été avancées pour expliquer le mécanisme du mouvement flagellaire. Butschli a observé que le flagelle subit une série de mouvements latéraux et, ce faisant, une pression est exercée sur l’eau perpendiculairement à sa surface.
Cette pression crée deux forces dirigées l’une parallèlement et l’autre perpendiculairement à l’axe principal du corps. La force parallèle poussera l’animal vers l’avant et la force agissant à angle droit ferait pivoter l’animal sur son propre axe.

Gray (1928) a suggéré qu’une série d’ondes passe d’une extrémité du flagelle à l’autre. Ces ondes créent deux types de forces, l’une dans la direction du mouvement et l’autre dans la direction circulaire avec l’axe principal du corps. Le premier fera avancer l’animal et le second fera tourner l’animal.
Pendant un temps assez long, on a généralement supposé que le flagelle était dirigé vers l’avant pendant le mouvement flagellaire, mais maintenant, il est généralement admis que le flagelle est droit et turgescent dans la course effective et est tombé vers l’arrière lors de la course de récupération.
Récemment, Lowndes (1941-43) a souligné que le flagelle est dirigé vers l’arrière pendant la locomotion. Selon Lowndes, une série d’ondes spirales passent successivement de la base à la pointe du flagelle dirigé vers l’arrière à environ 12 par seconde avec une vitesse et une amplitude croissantes.
Les vagues se déplacent le long du flagelle en spirale et font tourner le corps d’Euglena une fois par seconde. Ainsi, dans sa locomotion, il trace un chemin en spirale sur une ligne droite et avance. La vitesse de déplacement est de 3 mm par minute.
Cependant, le mouvement du flagelle est lié à la contraction de toutes ses fibrilles. L’énergie pour la contraction de ces fibrilles provient des ATP formés dans les mitochondries des blépharoplastes.
(ii) Mouvement euglénoïde ou Métaboly:
L’Euglène présente parfois des mouvements de tortillements lents très particuliers. Une vague péristaltique de contraction et d’expansion traverse tout le corps de l’extrémité antérieure à l’extrémité postérieure et l’animal avance. Le corps devient plus court et plus large d’abord à l’extrémité antérieure, puis au milieu et plus tard à l’extrémité postérieure.
Ce type de mouvement est appelé mouvement euglénoïde par lequel un mouvement lent et limité se produit. Les mouvements euglénoïdes sont provoqués par les contractions du cytoplasme ou par les contractions des myonèmes présents dans le cytoplasme sous la pellicule.
5. Nutrition d’Euglena Viridis:
Le mode de nutrition d’Euglena est mixotrophe, c’est-à-dire, la nutrition est accomplie soit par holophyte, soit par saprophyte, soit par les deux modes.
(i) Nutrition holophytique ou Autotrophe:
Chez Euglena, le principal mode de nutrition est holophytique ou de type végétal. L’aliment est fabriqué par photosynthèse, comme chez les plantes, à l’aide de dioxyde de carbone, de lumière et de chlorophylle présents dans les chromatophores. La chlorophylle décompose le dioxyde de carbone en carbone et en oxygène en présence de la lumière du soleil.
L’oxygène est libéré et le carbone est retenu et combiné avec les éléments de l’eau pour former des glucides (polysaccharides) comme le paramylum. Le paramyle diffère de l’amidon car il ne devient pas bleu avec une solution d’iode. Chez Euglena, la nourriture de réserve est stockée sous forme de corps paramylés réfractiles et leur nombre est abondant dans une Euglena bien nourrie.
(ii) Nutrition saprophyte ou Saprozoïque:
En l’absence de lumière du soleil, l’Euglène tire sa nourriture d’un autre mode de nutrition appelé saprophyte, osmotrophe ou saprozoïque. Dans ce mode, l’animal absorbe à travers sa surface corporelle générale des substances organiques en solution provenant de la matière en décomposition dans l’environnement de l’animal. Ils ont besoin de sels d’ammonium, au lieu de nitrates, pour leurs sources d’azote.
L’Euglène peut subsister grâce à la nutrition saprozoïque lorsqu’elle perd sa chlorophylle dans l’obscurité totale. Habituellement, les chlorophylles perdues dans l’obscurité sont retrouvées à la lumière. Mais dans des formes comme E. gracilis, le changement est permanent, c’est-à-dire que les chlorophylles une fois perdues ne sont pas retrouvées. La nutrition saprophyte peut également compléter la nutrition holophytique normale.
Une pinocytose a également été rapportée à la base du réservoir pour l’apport de protéines et d’autres grosses molécules. Lorsqu’un organisme présente en utilisant plus d’une méthode, on dit qu’il présente un mode de nutrition mixotrophe.
Euglena présente à la fois une nutrition holophytique et saprozoïque, il présente donc un mode de nutrition mixotrophe. La digestion est assurée par des enzymes sécrétées dans les vacuoles alimentaires par le cytoplasme environnant.
6. Respiration à Euglena Viridis:
Chez Euglena Viridis, l’échange de gaz (apport d’O2 et émission de CO2) se fait par diffusion à travers la surface du corps. Il absorbe l’oxygène dissous de l’eau environnante et dégage du dioxyde de carbone par diffusion.
Il y a tout lieu de croire que pendant la journée, l’oxygène libéré lors de la photosynthèse est utilisé à des fins de respiration et le dioxyde de carbone émis pendant la respiration peut être utilisé pour la photosynthèse.
7. Excrétion chez Euglena Viridis:
L’élimination du dioxyde de carbone et des déchets azotés (ammoniac) se fait à travers la surface générale du corps par diffusion. Au moins une certaine excrétion, cependant, est réalisée par la vacuole contractile.
Osmorégulation:
Depuis Euglena Viridis a une pellicule semi-perméable et vit dans l’eau de sorte que l’eau pénètre continuellement dans son corps par endosmose. L’élimination de l’excès d’eau du corps est connue sous le nom d’osmorégulation. L’élimination de l’excès d’eau se fait par la vacuole contractile.
Les vacuoles contractiles accessoires collectent l’excès d’eau du cytoplasme environnant et libèrent leur contenu dans la vacuole contractile principale qui augmente progressivement en taille et finit par éclater et forcer l’eau dans le réservoir. De l’eau du réservoir, s’échappe par le cytosome à travers le cytopharynx. Parallèlement à cela, les déchets solubles dans l’eau sont également jetés hors du corps.
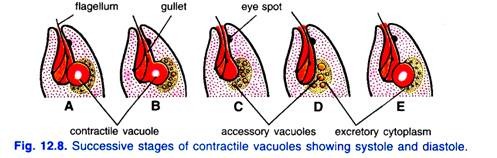
Récemment, Chadefaud a souligné que la vacuole contractile est entourée d’un cytoplasme granulaire et excréteur spécialisé. La vacuole contractile atteint périodiquement sa taille maximale et s’effondre pour rejeter son contenu dans le réservoir (c’est-à-dire la systole).
Simultanément, plusieurs petites vacuoles accessoires apparaissent dans le cytoplasme excréteur. Ces vacuoles fusionnent ensuite pour former une nouvelle grande vacuole (i.e., diastole) qui atteint la taille maximale et s’effondre pour évacuer l’eau comme la première.
8. Comportement d’Euglena Viridis:
Euglena Viridis répond à une variété de stimuli et est très.sensible à la lumière. Il nage vers une lumière ordinaire telle que celle d’une fenêtre et évite la lumière forte. Si une culture d’Euglena est examinée, la plupart des animaux se trouveront sur le côté vers la lumière. Ceci est d’un avantage certain pour l’animal, car la lumière est nécessaire à l’assimilation du dioxyde de carbone au moyen de sa chlorophylle.
Euglena nagera loin des rayons directs du soleil. La lumière directe du soleil tuera l’organisme si elle est autorisée à agir pendant une longue période. Si un plat contenant des Euglènes est placé à la lumière directe du soleil et que la moitié de celui-ci est ombragée, les animaux éviteront la partie ombragée ainsi que la lumière directe du soleil et resteront dans une petite bande entre les deux à la lumière qui leur convient le mieux (Fig. 12.9), c’est-à-dire leur optimum.
Une Euglène nageuse se déplace en spirale en tournant et en tournant autour de son propre axe, mais elle montre une réaction de choc chaque fois que la direction de la lumière est modifiée.

Il a été constaté que la région située devant la tache oculaire est plus sensible à la lumière que toute autre partie du corps. Euglena s’oriente parallèlement aux rayons de lumière chaque fois que le corps paraflagellaire (photorécepteur) est ombragé par le stigmate ou le trou oculaire. L’animal ajuste sa position à la direction de la lumière se déplaçant vers ou loin de lui.
Lorsque l’animal tourne, le stigmate agit comme un écran, le corps paraflagellaire est alternativement exposé ou protégé lorsque la lumière lui tombe sur le côté. L’animal s’ajuste jusqu’à ce que le corps paraflagellaire soit continuellement exposé, cela se produit lorsque la source de lumière est soit droite devant soit derrière.
Euglena permet d’éviter les réactions aux stimuli mécaniques, thermiques et chimiques selon un schéma d’essais et d’erreurs (phabotaxie). Lorsqu’elle est stimulée par un changement, l’Euglène, dans la majorité des cas, s’arrête ou recule, tourne fortement vers la surface dorsale, mais continue de tourner sur son grand axe.
L’extrémité postérieure fait alors office de pivot, tandis que l’extrémité antérieure trace un cercle de large diamètre dans l’eau. L’animal peut nager vers l’avant dans une nouvelle direction à partir de n’importe quel point de ce cercle. Cela évite la réaction.
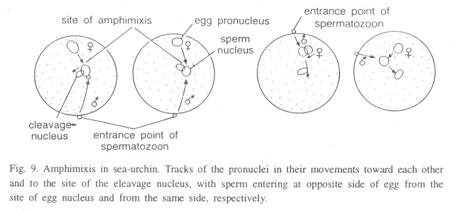
9. Reproduction chez Euglena Viridis :
Euglena Viridis se reproduit de manière asexuée par fission binaire longitudinale et fission multiple. L’enkystement a également lieu. La reproduction sexuée ne se produit pas, bien qu’une forme primitive de celle-ci soit signalée chez certaines espèces.
(i) Fission Binaire longitudinale:
Pendant les périodes actives, dans des conditions favorables d’eau, de température et de disponibilité alimentaire, Euglena se reproduit par fission binaire longitudinale. La fission est toujours symétrique, c’est-à-dire que l’Euglène parent se divise en deux euglènes filles, exactement identiques l’une à l’autre.
Le noyau se divise par mitose. L’endosome s’allonge transversalement et se rétrécit en deux parties approximativement égales. La division nucléaire a lieu à l’intérieur de la membrane nucléaire.
Les organites à l’extrémité antérieure tels que le stigmate, les blépharoplastes, le réservoir, le cytopharynx et les chromatophores et les corps paramylés sont également dupliqués. Le corps commence à se diviser dans le sens de la longueur, de l’extrémité antérieure vers le bas jusqu’à l’extrémité postérieure, ce qui entraîne la formation de deux individus filles.
L’ancien flagelle est retenu par une moitié, tandis qu’un nouveau flagelle est développé par l’autre, la vacuole contractile et le corps paraflagellaire ne se divisent pas mais disparaissent et se refont chez les individus filles.

( ii) Fission multiple:
La fission multiple a généralement lieu à l’état enkysté. Parfois, pendant les périodes de repos ou d’inactivité, l’enkystement se produit à Euglena. La masse du cytoplasme et le noyau à l’intérieur du kyste subissent des divisions mitotiques répétées donnant naissance à 16 ou 32 petites filles.
Au retour de conditions favorables, le kyste se brise et les individus filles s’échappent du kyste. Chaque individu fille développe les différents organites et commence la vie normale. Certains travailleurs considéraient les individus filles comme les spores et ce processus comme la sporulation.
(iii) Stade de la palmelle :
Parfois, généralement dans des conditions défavorables, un grand nombre d’euglenae se rapprochent, perdent leurs flagelles et deviennent arrondis. Ils sécrètent une matrice gélatineuse ou mucilagineuse à l’intérieur de laquelle ils restent incrustés. Cette condition est appelée stade de palmella qui est souvent considérée comme de l’écume verte à la surface de l’eau des étangs.
Les individus du stade palmella exercent des activités métaboliques et se reproduisent par fission binaire. À l’arrivée des conditions favorables, la couverture gélatineuse gonfle par l’absoprtion de l’eau et les euglènes sont libérées. Ils régénèrent leurs flagelles et commencent une vie active normale.
( iv) Enkystement:
Dans des conditions défavorables telles que la sécheresse, le froid extrême ou le chaud extrême, la pénurie de nourriture et d’oxygène, Euglena subit un enkystement. Tout d’abord, Euglena devient inactive, perd son flagelle et sécrète un kyste autour d’elle. Le kyste est sécrété par les corps mucifères situés sous la pellicule.
Le kyste est à paroi épaisse, arrondi et de couleur rouge en raison de la présence d’un pigment appelé hématochrome. Ce kyste est de type protecteur.
Pendant l’état enkysté, les périodes de conditions défavorables sont passées avec succès. Pendant l’enkystement, la fission binaire peut se produire une ou plusieurs fois, entraînant 2 à 32 petites euglènes filles dans le kyste. Au retour de conditions favorables, la paroi du kyste se brise, les animaux deviennent actifs et sortent du kyste pour mener une vie normale de nage libre.
En fait, l’enkystement ne se produit qu’à la marée au-dessus des conditions défavorables et, dans cette condition, la dispersion de l’Euglena se produit sur une vaste zone.
10. Position d’Euglena Viridis:
Euglena Viridis présente de nombreux caractères de plantes telles que les chloroplastes avec une nutrition chlorophylle et holophytique mais il est considéré comme un animal en raison des faits suivants:
(i) Sa pellicule est faite de protéines et non de cellulose comme chez les plantes.
(ii) Présence de blépharoplastes, comparables aux centrioles.
(iii) Présence du stigmate et du corps paraflagellaire, les structures photosensibles.
(iv) Présence de vacuoles contractiles, qui ne se trouvent pas chez les plantes.
(v) Mode de nutrition saprozoïque et aussi holozoïque comme l’ont affirmé certains zoologistes.
(vi) Présence de fission binaire longitudinale, que l’on ne trouve pas chez les plantes.
11. Quelques autres Flagellés Euglénoïdes :
(i) Euglena Gracilis:
Il est petit, allongé, en forme de fuseau, mesurant environ 50 microns de longueur. Les chloroplastes sont grands, plats, en forme de plaque et au nombre d’environ dix. Chaque chloroplaste porte un pyrénoïde protéiné. Contrairement à Euglena viridis, ses chloroplastes une fois perdus dans l’obscurité ne peuvent pas être récupérés. Son cytoplasme contient de nombreux corps paramylés en association avec les chloroplastes (Fig. 12.14 A).

( ii) Euglena Spirogyra:
Il est de grande taille et mesure environ 95 microns de longueur et 18 microns de largeur. Son corps est allongé, en forme de fuseau et en arrière son corps est étiré comme une queue. Il existe de nombreux petits chloroplastes en forme de disque sans pyrénoïdes. Il est caractérisé par les corps paramylés dans son cytoplasme (Fig. 12.14 B).
(iii) Astasia Longa:
C’est une forme euglénoïde typique et généralement supposée être la forme blanchie d’Euglena gracilis. Les chloroplastes, le stigmate et le corps paraflagellaire ne sont pas trouvés. Il présente une nutrition par osmotrophie due à l’absence de chloroplastes et son cytoplasme contient de nombreux corps paramylés (Fig. 12.14 C).
(iv) Paranema Trichophorum:
C’est un flagellé euglénoïde au corps quelque peu trapu. On pense qu’il se nourrit holozoïquement par phagotrophie sur des microorganismes assez gros. De ses deux flagelles, l’un est locomoteur et long, tandis que l’autre est traînant et se trouve attaché à la surface de son corps. Le stigmate et le corps paraflagellaire ne sont pas trouvés. Son cytoplasme contient des vacuoles alimentaires et de nombreux petits corps paramylés.
Il se caractérise par la présence d’un appareil accessoire en forme de bâtonnet appelé trichites dans son cytopharynx (Fig. 12.14 D).